

 PDF
PDF ePub
ePub Citation
Citation Print
Print


ABBREVIATIONS
NSC
neural stem cell
bHLH
basic helix-loop-helix
GFP
green fluorescent protein
SD
Sprague Dawley
EGF
epidermal growth factor
bFGF
basic fibroblast growth factor
PDL
poly-d-lysine
PBS
phosphate-buffered saline
DAPI
4',6-diamidino-2-phenylindole
ESC
embryonic stem cell
HSC
hematopoietic stem cell
AAV
adeno-associated virus
SCF
stem cell factor
INTRODUCTION
Stem cells are promising sources for cell therapy and drug screening because of their regeneration potential and pluri- or multi-potency [1-5]. Due to the therapeutic potential of stem cells, many researchers are intensively studying stem cells. To manipulate stem cells to express foreign genes to study the role of genes in stem cell biology, it is necessary to achieve high gene transduction efficiencies. Up-regulation or down-regulation of genes provides information about their roles in stem cell fate determination and is important to seek for the essential functions of genes [6-8]. However, stem cells are hard to be transfected with ectopic genes [9-13]. Among many types of stem cells, neural stem cells (NSCs) are multipotent cells that can proliferate and can give rise to different types of cells consisting of the nervous system such as neurons, astrocytes and oligodendrocytes [14]. Since NSCs can only differentiate into cells that compose the nervous system, they have been considered safe to be transplanted into body [3,15]. In many cases, NSCs have been transplanted into neurodegenerative conditions to replace degenerated neurons [2,3,16,17]. In order to elucidate the molecular mechanisms controlling NSCs fate, it is critical to achieve efficient gene delivery [18].
It has been previously reported that basic helix-loop-helix (bHLH) protein family including Ngn and ASCL1 play important roles in the generation of neurons [19,20]. During the development of the nervous system, generation of various types of neurons is temporally and spatially controlled. Neuronal differentiation may occur through direct transcriptional regulation by bHLH protein family [21-23]. To identify the functions of bHLH protein in stem cell, efficient gene delivery method has to be determined. Because NSCs are one of the hard-to-transfect cell types, we compared lipid mediated transfection, electroporation, nucleofection and viral transduction in NSCs derived from fetal rat cortex by using plasmid that encode green fluorescence protein (GFP). Furthermore, we observed Ngn2 introduced stem cell behavior using real time microscope and showed that over-expression of Ngn2, a bHLH protein family, leads to an increased final cell division. The development of an efficient gene delivery method in NSCs may be useful in determining functions of genes in stem cells and used to develop stem cell based drug screening system.
Go to : 
METHODS
Rat NSC cultures
The cortex of E14 Sprague Dawley (SD) rat brain was isolated and dissociated with accutase (Millipore, USA). Starting from 200,000/ml density, cells were expanded as neurospheres in Dulbecco's Modified Eagle Medium: Nutrient Mixture F-12 (DMEM/F12, Invitrogen, USA) containing 1% antibiotic-antimycotic (v/v) (Invitrogen, USA), 2% B27 (v/v) (Invitrogen, USA), 20 ng/ml epidermal growth factor (EGF, Millipore, USA) and 20 ng/ml basic fibroblast growth factor (bFGF, Millipore, USA). The medium was conditionally changed every other day by removing half the media and adding fresh media containing 2% B27 and 20 ng/ml EGF and bFGF. After 1 week, cells were dissociated with accutase and used for lipid-mediated transfection, electroporation, nucleofection, and retrovirus transduction.
Lipid-mediated transfection
Lipid-mediated transfection was performed using X-tremeGENE 9 DNA Transfection Reagent (Roche, Germany) according to the manufacturer's protocol. After plating 6×104 cells at the poly-d-lysine (PDL, Sigma-Aldrich, USA) and laminin (Invitrogen, USA) coated microscope cover glasses (Superior, Germany) in 24 well plates (Nunc, Germany), NSCs were cultured for 1 day to 50~60% confluency and washed twice with DMEM/F12 and then the media was changed to Opti-MEM (Invitrogen, USA). Each well was added with 250 ng retroviral GFP vector, 0.75 µl X-treme-GENE 9 DNA Transfection Reagent and 25 µl Opti-MEM mixture. After 8 hours incubation, cells were washed twice with DMEM/F12 and then medium was changed to proliferation media containing 2% B27 and 20 ng/ml EGF and bFGF. Two days after the transfection, cells were fixed for immunocytochemistry.
Electroporation
Electroporation was performed using NEPA21 electroporator (Nepagene, Japan) according to the manufacturer's recommendation. 1×106 dissociated rat NSCs were centrifuged at 90 g for 10 minutes at room temperature and resuspended in 100 µl mixture of Opti-MEM and 10 µg retroviral GFP vector. The mixture received two types of pulses, poring pulse and transfer pulse. Poring pulse condition was as below: 150 V, 5 miliseconds (ms) pulse length, total two pulses, 50 ms interval between the pulses, 10% decay (D). Rate with + polarity. The transfer pulse condition was as below: 20 V, 50 ms pulse length, total five pulses, 50 ms interval among the pulses, 40% D. Rate with +/- polarity. After the electroporation, cells were immediately seeded onto PDL and laminin coated microscope cover glasses in 24 well plates. Two days after the electroporation, cells were fixed for immunocytochemistry.
Nucleofection
Nucleofection was performed using Amaxa 4D nucleofector (Lonza, Switzerland) using P3 and P4 Primary Cell 4D-Nucleofector® X Kit L (Lonza, Switzerland) according to the manufacturer's protocol. 5×106 dissociated rat NSCs were centrifuged at 90 g for 10 minutes at room temperature and resuspended in 100 µl nucleofector solution. 5 µg of retroviral GFP vector was added and the mixture was transferred to 100 µl Single Nucleocuvette. After the nucleofection, cells were immediately seeded onto PDL and laminin coated microscope cover glasses in 24 well plates. Two days after the nucleofection, cells were fixed for immunocytochemistry.
Retrovirus preparation and transduction
Retroviral GFP vector and retroviral Ngn2 vector were used to generate retroviruses. PLAT-E cells transduced with gag-pol and env were used for the packaging of viruses. Briefly, PLAT-E cells were transfected with retroviral GFP vector or Ngn2 vector using X-tremeGENE 9 following manufacturer's instruction. Supernatant containing viral particles was collected 2 days after transfection and centrifuged at 12,500 g for 12 hours to concentrate retroviruses. NSCs were plated at the PDL/laminin-coated microscope cover glasses in 24 well plates and cultured for 1 day to 50~60% confluency, and then NSCs were exposed to retroviruses for 6 hours in the presence of polybrene (4 µg/ml, Sigma-Aldrich, USA). After washing twice with DMEM/F12, cells were cultured in the presence of bFGF and EGF for 2 days. Then NSCs were fixed for immunocytochemistry.
Immunocytochemistry and cell counting
NSCs were fixed for 30 minutes with 4% paraformaldehyde (Affymetrix, USA) and rinsed twice with phosphate-buffered saline (PBS). Fixed cells were blocked for 30 minutes in PBS containing 5% normal goat serum (Millipore, USA) and 0.2% triton X-100 (Amresco, USA) and incubated 1 hour with anti-GFP primary antibody (1:500, Molecular Probes, USA). Following rinsing with PBS, cells were incubated with Alexa Fluor 488-conjugated secondary antibody (1:1,000, Molecular Probes, USA). Following rinsing with PBS, cell nuclei were stained by 4'-6-Diamidino-2-phenylindole (DAPI) (0.5 µg/ml in PBS, Sigma-Aldrich, USA) to count the total cell number. The images were obtained with an inverse fluorescence microscopy (DMIL; Leica, Germany). Cell counting was performed in 3 randomly chosen microscopic fields. The ratio of GFP-positive cells to total cells was calculated to draw transfection or transduction efficiency.
Time-lapse analysis
Inverted Nikon microscope (Nikon, Japan) was used to capture images for GFP-positive cells. Fluorescence images were acquired every 4 minutes to trace behaviors of the GFP-positive cells. Cells were kept in humidified, 37℃ temperature control chamber with 4~6% CO2 during the image recording. Acquired images were assembled into movies using Metamorph imaging system (Molecular Devices, USA). The movies were used to evaluate the behavior and cell division of individual cells.
Statistics
Differences between groups were tested for significance using Student's t-test and considered to be statistically significant if p<0.05. Quantitative data were expressed as mean±S.D.
Go to :
RESULTS
Lipid-mediated transfection showed low transfection efficiency in NSCs
Cell therapy using stem cells and understanding the mechanisms how stem cells proliferate and differentiate require safe, efficient and stable delivery of foreign genes without making any harmful effect and changing essential properties. Lipid-mediated transfection is one of the most widely used gene delivery methods. To explore the efficiency of lipid mediated transfection in rat NSCs, we used retroviral vector encoding GFP. Cortical NSCs were cultured in the presence of EGF and bFGF for 1 week, then dissociated and plated onto PDL and laminin coated coverslips. One day after plating, NSCs were transfected with X-tremeGENE9 alone or with X-tremeGENE 9+retroviral GFP vector. Two days after transfection, NSCs were fixed and immunostained with anti-GFP antibody to determine the transfection efficiency. To count the total cell number, nuclei were stained with DAPI. As shown in Fig. 1, the efficiency of lipid-mediated transfection in rat NSCs was only around 4.48%. These data suggest that lipid mediated transfection is not effective in NSCs for ectopic expression of foreign genes.
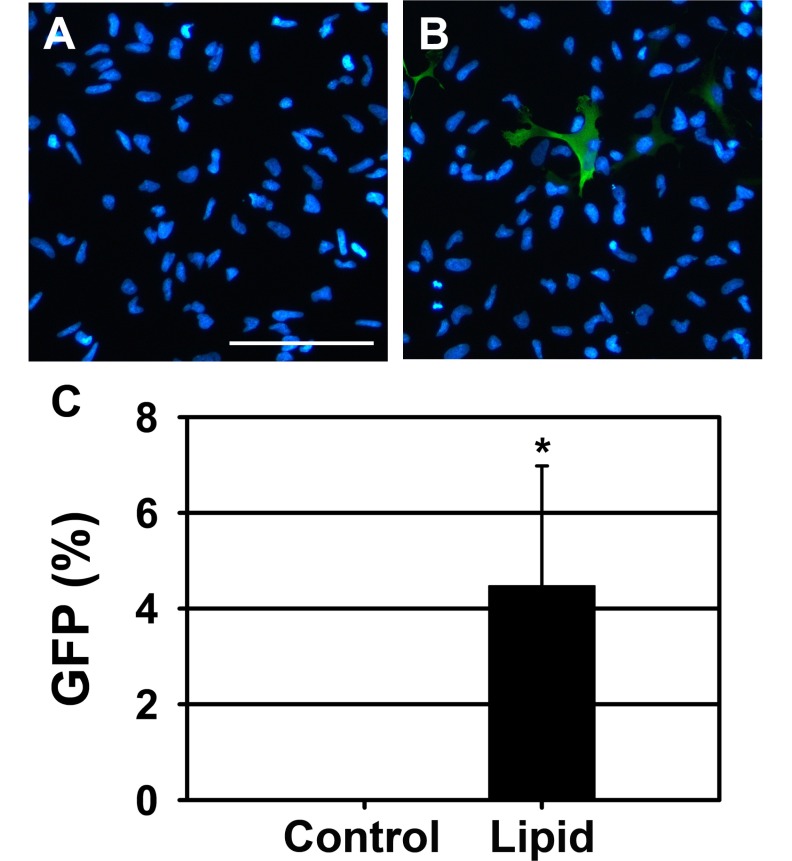 | Fig. 1Transfection efficiency of rat NSCs 48 hours after lipid-mediated transfection. (A, B) Representative merged images of NSCs transfected with (A) lipid alone and (B) lipid with GFP vector. GFP was visualized by immunostaining with anti-GFP primary antibody and Alexa Fluor 488-conjugated secondary antibody (green). Nuclei were visualized by DAPI staining (blue). Scale bar=100 µm. (C) Quantification of the immunostaining data. The ratio of GFP-positive cells to the total cells was calculated and presented. The data are shown as mean±S.D. (*p<0.05 vs. negative control).
|
Nucleofection and retrovirus transduction showed better transfection efficiency
Since lipid-mediated transfection efficiency was only around 3~4% (Fig. 1B, 1C and 2A), we next decided to compare transfection efficiencies of electroporation, nucleofection and retrovirus transduction in NSCs. NSCs were expanded as neurospheres for a week in the presence of EGF and bFGF. Then NSCs were dissociated to make single cell suspension and directly used for electroporation or nucleofection. In the electroporation experiments using NEPA21 electroporator, 1×106 cells were mixed with 10 µg of retroviral GFP vector in 100 µl Opti-MEM and then electroporated using following conditions. NSCs and GFP vector were mixed and electroporated twice with 150 V pulse for 5 ms and between the pulses 50 ms interval was given (this is to make pores in the NSCs). Then, additional five times of 20 V pulse for 50 ms with 50 ms intervals among the pulses (this is to deliver GFP vector to pored NSCs). After the electroporation, cells were directly transferred to pre-warmed medium containing EGF, bFGF and 2 µg/ml laminin and grown as monolayer for 2 days. Then NSCs were fixed and immunostained with anti GFP antibody. Using electroporation, the transfection efficiency was improved to 22.72±1.48% (Fig. 2B and 2E) which was seven times higher than lipid-mediated transfection.
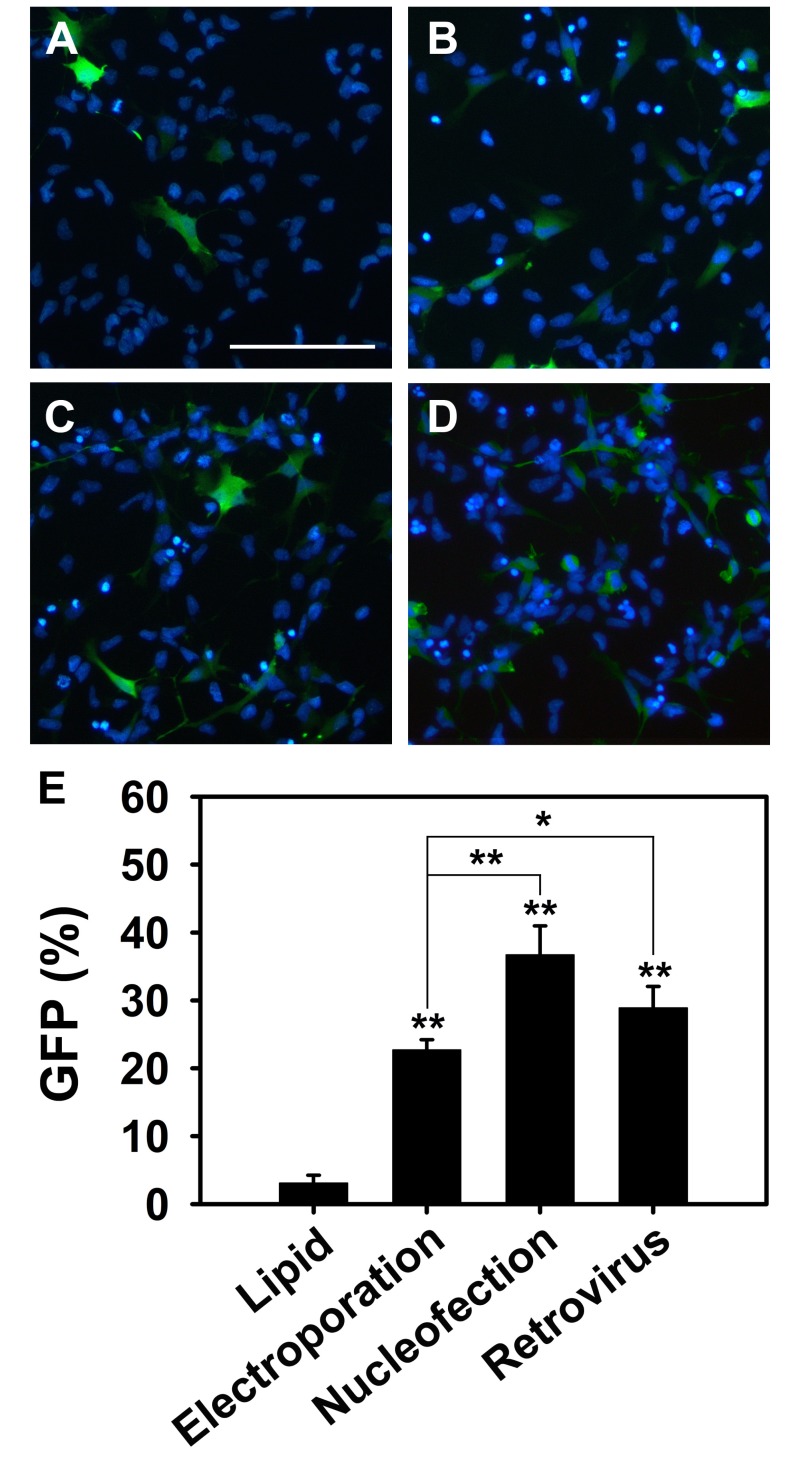 | Fig. 2Comparison of transfection efficiency of lipid-mediated transfection, electroporation, nucleofection and retroviral transduction. (A~D) Representative merged images of NSCs transfected with GFP vector for 48 hours using (A) X-tremeGENE9 transfection reagent, (B) NEPA21 electroporator, (C) Amaxa 4D Nucleofector, and (D) retroviral transduction. By immunostaining GFP positive cells are shown in green and nuclei are shown in blue. Scale bar=100 µm. (E) Quantification data of the transfected cells. The ratio of GFP-positive cells to the total cells was calculated. Quantitative data are shown as mean±S.D. (**p<0.01, *p<0.05).
|
Nucleofection is also an electroporation-based technique, however, it aims to transfer DNA or RNA directly into the cell nucleus. To transfect NSCs by nucleofection, 5×106 cells were resuspended in 100 µl mixture of solution provided by the manufacturer (P4 Primary Cell 4D-Nucleofector® X Kit) and 5 µg of retroviral GFP vector DNA was added. After the nucleofection using CA137, CL133, CU110, DC100, DC104, DR114 and EM110 programs, cells were plated into 24 well plates and grown as monolayer for additional 2 days in serum free media with EGF and bFGF. Among the 7 programs, CA137 showed the highest transfection efficiency (36.71±4.29%) in rat NSCs (Fig. 2C and 2E).
To explore gene delivery efficiency of retroviral transduction, NSCs were dissociated and 6×104 cells were plated onto PDL and laminin coated 24 well plates. One day after plating, NSCs were transduced with retroviruses (1 MOI) encoding the GFP and cultured in media with EGF and bFGF. Two days after retroviral transduction, NSCs were fixed and immunostained with anti-GFP antibody. Around 30% (28.90±3.16%) of total cells were expressing GFP (Fig. 2D). These data show that electroporation, nucleofection and retroviral transduction are more effective to introduce ectopic genes to NSCs than lipid-mediated transfection. In addition, among the conditions that we tested, nucleofection and retroviral transduction significantly increased the numbers of transfected NSCs compared to electroporation.
Recently, Bertram and colleagues reported that mouse NSCs showed more than 80% transfection efficiency when they used DS112 and DS113 nucleofection programs and P3 primary cell 4D-nucleofector® X kit solution [18]. Therefore, we compared transfection efficiency of DS112, DS113 using P3 primary cell 4D-nucleofector® X kit solution and CA137 using P4 primary cell 4D-nucleofector® X kit solution in rat NSCs. Surprisingly, DS112 (4.71±0.39%) and DS113 (8.38±1.91%) showed far lower transfection efficiency than CA137 (27.96±2.53%) (Fig. 3). These results indicate that for rat NSCs transfection, using CA137 program with P4 solution is more efficient than using DS112 and DS113 with P3 solution.
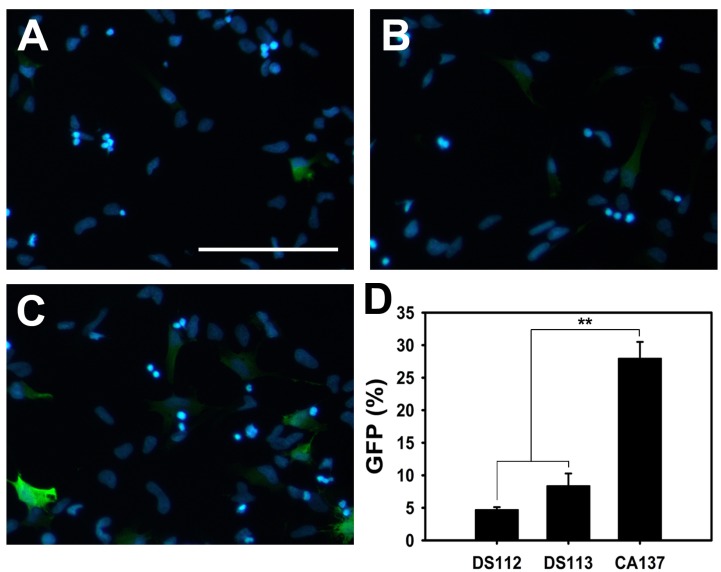 | Fig. 3Comparison of transfection efficiency of three programs -DS112, DS113 and CA137- of nucleofection. (A~C) Representative merged images of NSCs nucleofected using program (A), (B) DS113 and (C) CA137 for 48 hours. GFP positive cells are shown in green and DAPI stained nuclei are shown in blue. Scale bar=100 µm. (D) Quantification of the transfected NSCs. The ratio of GFP-positive cells to the total cells was calculated and presented. Quantitative data are shown as mean±S.D. (**p<0.01).
|
Ectopic expression of Ngn2 facilitates final cell division
Our data showed that electroporation, nucleofection and retroviral transduction provides above 20% gene delivery in NSCs. To explore transfected gene functions in NSCs, we next overexpressed Ngn2 that are known to be involved in neurogenesis [24]. After retroviral transduction of Ngn2 in NSCs, time-lapse analysis revealed enhanced cell division. About 8 hours after viral transduction, NSCs started to express ectopic genes that are visualized by GFP. Time-lapse microscopy data showed 1.3 times higher cell division in NSCs expressing Ngn2 than those expressing GFP alone (Fig. 4). These data suggest that Ngn2 promotes final cell division before differentiate into neuron. Our data proves the concept that ectopic genes function in NSCs using retroviral transduction.
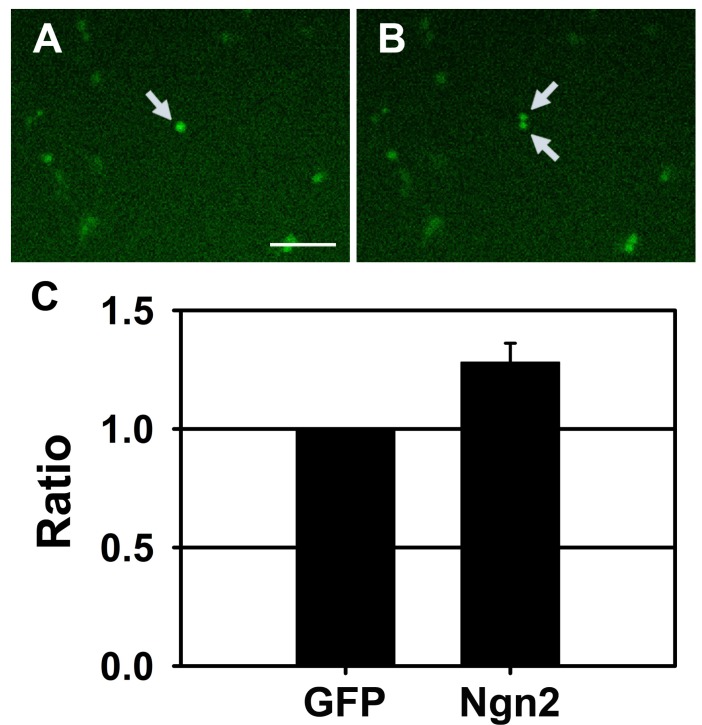 | Fig. 4Induction of final cell divisions by Ngn2. (A, B) Representative fluorescence time-lapse microscopic images of NSCs transduced with Ngn2 retrovirus. (A) Arrow-pointed cell rounded up and divided into (B) two daughter cells. Scale bar=100 µm (C) Quantification of cell division observed in NSCs transduced with control GFP retrovirus or Ngn2 retrovirus. Time-lapse video was recorded for about 24 hours. The ratio of dividing GFP-positive cells to the total GFP-positive cells was calculated and presented. Quantitative data are shown as mean±S.D.
|
Go to :
DISCUSSION
Until recently, ectopic genes were introduced into various types of mammalian cells by the methods using calcium phosphate, polycations and liposome [25]. Among those, lipid-mediated transfection has been most frequently used for overexpression in various cell types [26,27]. However, some of cells including embryonic stem cells (ESCs) and NSCs are not effectively transfected by lipid mediated methods [9,11]. It is reported that with lipofection, about 10.7% of transfection efficiency has been achieved [10,12,13]. However, our study with lipid mediated transfection, we could only achieve around 3~4% of transfection efficiency. If we consider the fact that after NSCs differentiation, different types of cells including neurons, astrocytes, oligodendrocytes and undifferentiated NSCs exist in a culture well, with such low efficiency, it is hard to determine the effects of transgenes. Thus several alternative methods have been addressed to enhance ectopic gene expression. Viral transduction using retrovirus, lentivirus or adenovirus, electroporation and nucleofection provide better efficiencies [12,18,28-33].
In this study, when compared with lipid-mediated transfection, electroporation or nucleofection significantly improved transfection efficiency. With both methods, we could observe around 20~30% of GFP positive cells. Electroporation is widely used for in vitro and in vivo transfection and utilizes high voltage to permeabilize cell membrane to transport DNA into the cells [29-31,33]. Nucleofection is an electroporation-based technology and has been shown to introduce ectopic genes in various types of cells including stem cells and primary cells [32,34-36]. Nucleofection is preferentially used to deliver ectopic genes into hard-to-transfect stem cells such as ESCs, messenchymal stem cells, hematopoietic stem cells (HSCs) and NSCs due to high gene transfer efficiency [11,12,18,28,37-39]. Nucleofection showed better transfection efficiency over electroporation in human ESCs (hESCs) and rodent NSCs, probably because DNA was able to reach not only in the cytoplasm but also in the nucleus [12]. Similar to this, in our experiment, nucleofection showed higher transfection efficiency than electroporation. However, it is still possible that the optimal conditions for electroporation have not been determined yet. A recent study by Bertram and colleagues showed 88% transfection efficiency in mouse fetal cortical NSCs using amaxa 4D nucleofector program DS113 [18]. However, using the same protocol, we could only detect 4.71±0.39% of transfected rat NSCs with DS112, and 8.38±1.91% with DS113. In addition, the program CA137 was three times more efficient for the transfection of rat NSCs. The difference between our data and theirs could be from the different origin of NSCs (mouse or rat), or different GFP expression vectors that were used to trace transfected cells.
Viral transduction is another method to introduce foreign genes into cells [40-43]. In our experiments, gene delivery efficiency using retrovirus was comparable to that of nucleofection (around 30%). Although the transfection efficiency can be improved by using high titered virus, with 1MOI virus, we usually achieved around 30% transduction. With retrovirus transduction ectopic gene functions have been identified in NSCs [24,44]. We showed that the overexpression of bHLH proteins using retroviral transduction significantly induced neurogenesis in NSCs [24]. Lu and colleagues also showed that genetic modification with neurotrophin-3 using retroviruses promoted the survival, proliferation, neuronal differentiation and elongation of neurites in human NSCs [44]. It has been previously reported higher efficiencies can be achieved using other types of virus in different types of cells [1,45-47]. With adeno-associated virus (AAV), it has been reported that up to 50% of hESCs can be transfected [47]. Similarly, adenovirus mediated transfection also showed about 50% of efficiency in adult rat subventricular zone-derived NSCs [46]. With lentivirus, the transfection efficiency goes up much higher, almost 80% of human and rat NSCs can express transgenes [1,45]. Because only cells that are replicating at the time of infection can be transduced by retrovirus, transduction using retrovirus shows lower transduction efficiency compared to AAV, adenovirus or lentivirus [48,49]. Retroviral transduction requires breakdown of the nuclear envelope that occurs during mitosis [49-51]. In contrast, AAV, adenovirus and lentivirus can infect terminally differentiated, non-dividing cells as well as dividing cells [52,53]. However, one of the advantage of using retrovirus is that since they only infect cells that are dividing at the time of transduction, terminally differentiated cells can be excluded and only multipotent or pluripotent stem cells can be marked by the transgene.
To improve retroviral transduction efficiency, supplement of growth factor has been used to boost up the mitosis. It is reported that the use of stem cell factor (SCF) in combination with interleukin-6 in murine HSCs improved retroviral transduction efficiency [54]. Granulocyte-colony stimulating factor/SCF or Flk-2/Flt3 ligand/interleukin-3 are also suggested to improve retroviral transduction efficiency in HSCs [55,56]. Even in the lentivirus mediated transduction, it has been known that EGF or hepatocyte growth factor markedly improved gene transfer [1,57]. Those factors may provide protective effects onto cells to improve transduction efficiency in viral transduction. In addition, to improve viral transduction efficiency, magnetically guided AAV delivery system was used and this significantly enhanced cellular transduction in human NSCs (hNSCs) and reduced the virus incubation time [58]. Elastin-like polypeptide-treated polystyrene surfaces are known to enhance AAV-mediated gene delivery efficiency in hNSCs [59]. It is also reported that ultrasound can enhance retrovirus-mediated gene transfer efficiency in various cell types [60,61]. It is suggested that exposure times of virus or timing of infection could affect viral transduction efficiency. However, the optimum time of virus exposure to cells may vary depending on cell types [20,46].
Cationic agents such as polybrene are generally used for retroviral-mediated gene transfer because they have been known to increase the efficiency of retroviral transduction [62-64]. Both the lipid membranes of the cell and viral particles have net negative charges. Cationic polymers are capable of overcoming this electrostatic repulsion via neutralization of negative cells and virus surface charges, and thus enhance retroviral transduction efficiency [64-66]. Since cationic polymers show their role in enhancement of transduction efficiency via charge neutralization, anionic polymers are considered to have the reverse effect on transduction efficiency and some studies showed that anionic polymers inhibit the transduction processes [66-68]. However, other studies showed opposite results that retroviral transduction efficiency is enhanced when an anionic polymer is added with equal or higher weight concentration of a cationic polymer [69,70]. Recently, several studies have demonstrated that some other cationic agents including cationic liposomes can increase retroviral transduction efficiency compared to polybrene [62,63,71,72]. In our study, we have used polybrene to increase the transduction efficiency. Without polybrene, we could observe only around 3% transduction efficiency (data not shown). It is considered that enhancing effects of retroviral transduction efficiency by cationic polymers depends on both the type of target cells and envelope proteins used for pseudotyping [73]. Thus we can achieve better retroviral transduction efficiencies in NSCs by choosing an appropriate cationic agent.
Several earlier studies reported that ectopic expression of bHLH genes like Ngn2 can induce the generation of neurons from different types of cells [20,24,74-76]. In our current study, we observed facilitated cell division from NSCs that are retrovirally tranduced with Ngn2. It has been known that neurons are born by both asymmetric and symmetric divisions [77]. The induction of cell divisions in Ngn2 expressing cells shown by time-lapse analysis in our study indicates that Ngn2 may facilitate final symmetric cell divisions and produce two daughter neurons. This observation is consistent with the previous findings that Ngn2 infected colonies of neural crest stem cells or rat NSCs were smaller and contained a higher percentage of neurons than ASCL1 infected colonies [20,24,78].
Recently, a new transfection technology has been introduced. Pickard and colleagues reported that they could efficiently transfect mouse NSCs with using magnetofection, a technique that utilizes magnetic nanoparticles for gene delivery [79]. Thus, it will be interesting to compare viral infection, nucleofection and magnetofection in terms of transfection efficiency, cell viability and toxicity.
In this study, we have compared the transfection efficiency of lipofection, electroporation, nucleofection and retroviral transduction using rat NSCs and demonstrated that the nucleofection and retroviral transduction were better than lipid mediated transfection and electroporation. Furthermore, we have demonstrated that ectopic expression of Ngn2 can induce final cell division for differentiation indicating using these techniques, the functions of genes can be explored in stem cells.
Go to :
 XML Download
XML Download