

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
γ-Aminobutyric acid (GABA) receptors are members of the Cys-loop family with ligand-gated ion channels, and the Cys-loop family includes proteins such as 5-hydroxytryptamine 3 (5-HT3), nicotinic acetylcholine, and glycine receptors [1]. The structure of the ligand-gated ion channels is similar to that of the pentameric ion channels. In addition, the ion channels make channel pores through the transmembrane 2 (TM2) domain [1]. The GABA receptors are classified into 3 types of receptors: GABAA, GABAB, and GABAC. The GABAA and GABAC receptors function as anion selective channels that are permeable to chloride ions, whereas GABAB receptors are G-protein-coupled receptors [2,3]. GABAA receptors are composed of heteropentameric anion channels composed of α, β, γ, δ, ε, θ and π subunits; however GABAC receptors can form homopentameric or heteropentameric anion channels by ρ subunits alone or with α1 and γ2 [2-4]. The GABAA receptor is predominantly expressed in the central nervous system [5,6], whereas GABAC receptors are mainly expressed in retinal bipolar cells [7,8] and have a lower abundance in cerebellum [9] and hippocampus [10]. They play an important role in vision, sleep, cognition, and memory [11].
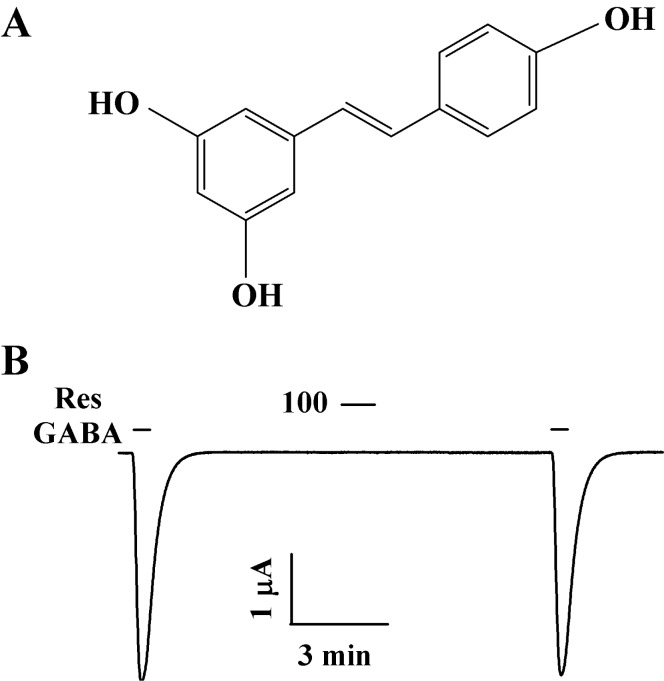
Resveratrol is a phytoalexin found in grapes, red wine, and other berries (Fig. 1A) and is also produced as an anti-fungal chemical by plants [12]. The concentration of resveratrol in red wine is as high as 0.2~5.8 mg/l [13]. Resveratrol exhibits diverse physiological and pharmacological activities, such as anti-cancer, chemopreventive, antiviral, cardio-protective, anti-aging, anti-inflammatory, and life-prolonging effects [13-15]. Resveratrol also has neuroprotective effects, and it attenuates neurodegenerative disorders such as Alzheimer's disease [16]. It also attenuates neuronal cell death caused by in vitro or in vivo brain hypoxia or ischemic conditions [17,18]. Although accumulating evidence indicates that resveratrol has diverse beneficial properties, including protective effects on the nervous systems, relatively little is known about its effects on cells, especially with respect to the regulation of receptors involved in synaptic transmission.
Recently, we demonstrated that resveratrol regulates 5-HT3A receptor expression [19]. In the present study, we examined the effects of resveratrol on homomeric GABAC receptor channel activity and found that resveratrol inhibits GABAC receptor channel activity in a concentration-dependent, voltage-independent, and non-competitive manner. These results indicate that resveratrol might play a role in the regulation of GABAC receptor channel activities.
Go to : 
METHODS
Materials
The cDNAs for human GABAC receptor ρ1 subunit were purchased from Thermo Fisher Scientific Inc. (Wyman Street Waltham, MA, USA). Fig. 1A shows the structure of resveratrol. Resveratrol used in this study was dissolved in dimethyl sulfoxide (DMSO) as previously reported [20] and was diluted with bath medium before use. Resveratrol was stored in the dark because it is light sensitive, and a fresh stock solution was prepared for every experiment. The final DMSO concentration was less than 0.1%. Resveratrol and other chemicals were purchased from Sigma-Aldrich (St. Louis, MO, USA).
Oocyte preparation
Xenopus laevis care and handling were performed in accordance to the guide for the Care and Use of Laboratory Animals, published by NIH, USA. Frogs underwent surgery only twice, separated by an interval of at least 3 weeks. Frogs were anesthetized with an aerated solution of 3-aminobenzoic acid ethyl ester for oocyte isolation. Oocytes were separated by collagenase treatment along with gentle shaking for 2 h in a CaCl2-free medium containing 82.5 mM NaCl, 2 mM KCl, 1 mM MgCl2, 5 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES), 2.5 mM sodium pyruvate, 100 units/ml penicillin, and 100 µg/ml streptomycin. Only stage 5 or 6 oocytes were collected and maintained at 18℃, with continuous gentle shaking in ND96 (96 mM NaCl, 2 mM KCl, 1 mM MgCl2, 1.8 mM CaCl2, and 5 mM HEPES; pH 7.5) supplemented with 0.5 mM theophylline and 50 µg/ml gentamycin. All the solutions were changed daily. All the experiments were performed within 2~4 days following the isolation of the oocytes [21].
Oocyte recording
A single oocyte was placed in a small Plexiglas net chamber (0.5 ml) and was constantly superfused with ND96 medium in the presence or absence of GABA or resveratrol during recording. The microelectrodes were filled with 3 M KCl and had a resistance of 0.2~0.7 MΩ. Two-electrode voltage-clamp recordings were performed at room temperature using Oocyte Clamp (OC-725C, Warner Instrument) with Digidata 1200A. For most of the electrophysiological experiments, the oocytes were clamped at a holding potential of -80 mV. For the current-voltage relationship, voltage ramps were applied from -100 to +40 mV for 300 ms.
cRNA preparation of GABAC receptor ρ1 and microinjection
A recombinant plasmid containing a human GABAC receptor ρ1 cDNA insert was linearized by digestion with the appropriate restriction enzymes. The cRNAs from linearized templates were obtained by using an in vitro transcription kit (mMessage mMachine; Ambion, Austin, TX) with a T3 polymerase. The RNA was dissolved in RNase-free water at 1 µg/µl, divided into aliquots, and stored at -80℃ until use. Oocytes were injected with H2O or mouse glycine α1 receptor cRNAs (5~10 ng) by using a Nanoject Automatic Oocyte Injector (Drummond Scientific, Broomall, PA). The injection pipette was pulled from the glass capillary tubing used for the recording electrodes, and the tip was broken to obtain an outer diameter of approximately 20 µm [21]. The final cRNA products were resuspended with RNase-free water at a concentration of 1 µg/µl and stored at -80℃.
Data analysis
To obtain the concentration-response curve for GABA-induced current in the presence of resveratrol, the observed peak amplitudes were normalized and plotted, and then fitted to the Hill equation, described below, by using Origin software (Northampton, MA): y/ymax=[A]n/([A]n+[IC50]n), where y represents percent (%) inhibition at a given concentration of resveratrol, ymax represents percent (%) maximal inhibition, IC50 is the concentration of resveratrol producing half-maximum inhibition of the control response to GABA, [A] is the concentration of resveratrol, and n is the interaction coefficient. All the values are presented as the mean±S.E.M. The differences between mean values of control and that of resveratrol treatment data were analyzed using unpaired Student's t test and one-way ANOVA test. A value of p<0.05 was considered statistically significant.
Go to :
RESULTS
Effect of resveratrol on IGABA in oocytes that express homomeric GABAC receptors
The addition of GABA to the bathing solution induced a large inward current in oocytes injected with GABAC receptor, indicating that functional GABAC receptors were expressed by the oocytes (Fig. 1B). Resveratrol itself had no effect on oocytes with GABAC receptors at a holding potential of -80 mV (Fig. 1B). Interestingly, pre-treatment of resveratrol induced a much larger inhibition of IGABA than that after co-treatment (Fig. 2A and B, n=9 from 3 frogs). The inhibition of IGABA by resveratrol in oocytes with GABAC receptors was reversible (Fig. 2A). Thus, these results show that resveratrol may regulate GABAC receptors channel activity, although resveratrol itself had no effect on GABAC receptor channel activity.
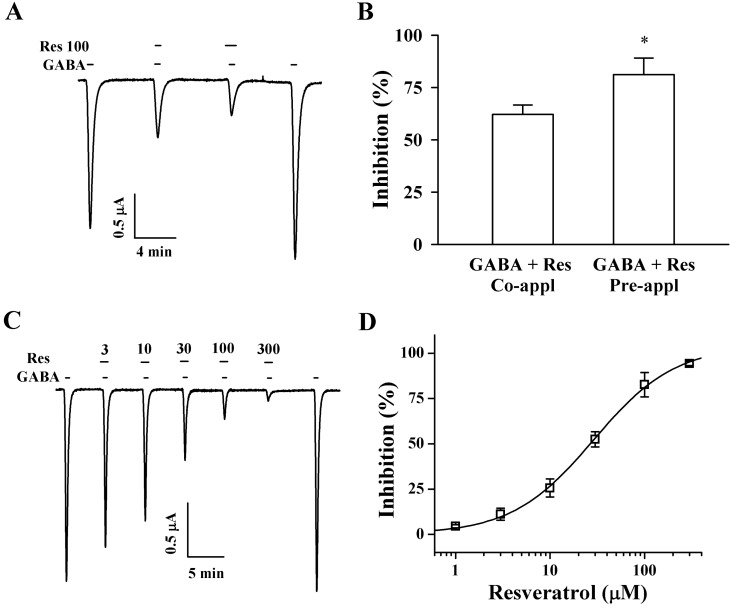 | Fig. 2Effect of Res on IGABA expression in oocytes that express GABAC receptors. (A) GABA (2 µM) was first applied and then GABA was co- or pre-applied with Res (100 µM). Thus, co- and pre-application of Res with GABA inhibited IGABA. The resting membrane potential of oocytes was approximately -35 mV, and oocytes were voltage clamped at a holding potential of -80 mV prior to drug application. Traces are representative of 6 separate oocytes from 3 frogs. (B) Pre-application of Res inhibited IGABA more potently than that inhibited by co-treatment. (C) IGABA in GABAC receptors expressing oocytes was elicited at -80 mV holding potential, with indicated time in the presence of 2 µM GABA and with the indicated pre-treatment concentration of Res that was applied before GABA application. (D) IGABA % inhibition induced by Res treatment was calculated using the average of peak that the inward current elicited by GABA treatment before Res application and of the peak inward current elicited by GABA treatment after pre-treatment of Res before GABA. The continuous line shows the curve fitted according to the equation. Each point represents the mean±S.E.M. (n=9~12 from 3 frogs).
|
Concentration-dependent effect of resveratrol on IGABA in oocytes with GABAC receptor
Since pre-treatment of resveratrol enhanced the inhibition on IGABA in oocytes that expressed GABAC receptor as compared to the IGABA inhibition in co-treated oocytes, we examined the effects of resveratrol on IGABA after the pre-treatment of resveratrol with GABA. In concentration-response experiments, the pre-treatment of resveratrol with GABA inhibited IGABA in a concentration-dependent manner in oocytes with GABAC receptor (Fig. 2C). The IC50 of IGABA was 28.9±2.8 µM for oocytes that expressed GABAC receptors (n=9~12 from 3 frogs) (Fig. 2D). Since the pre-application of resveratrol showed a stronger decrease of IGABA than that observed for the co-application of resveratrol, we examined the time-dependent effects of resveratrol preapplication. As shown in Fig. 3A, the pre-application of resveratrol enhanced IGABA inhibition and was time-dependent, while the time-dependent effects of resveratrol were almost saturated at 30 sec pre-application (Fig. 3B).
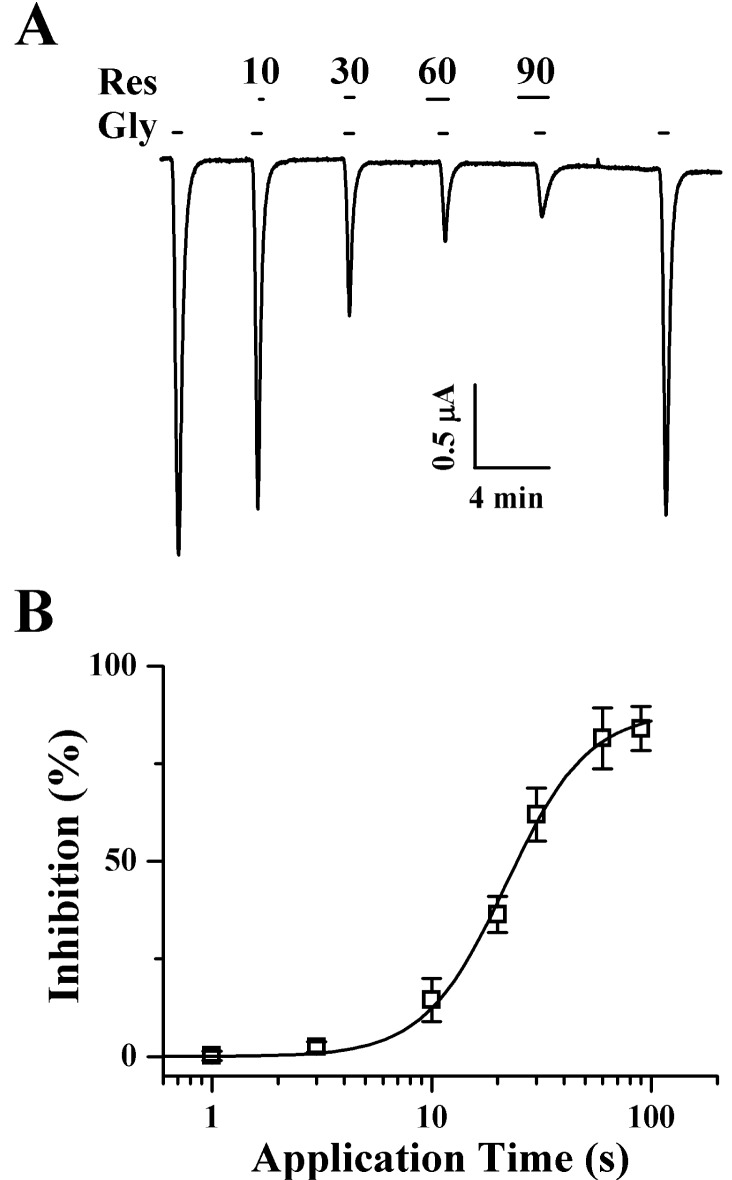 | Fig. 3Time-dependent effects of pre-application of Res on IGABA in oocytes that express GABAC receptors. (A) Res (100 µM)-mediated inhibition on IGABA is pre-application-time dependent. Traces represent 6 separate oocytes from 3 frog batches. IGABA in GABAC receptor-expressing oocytes was elicited at a holding potential of -80 mV for the indicated Res pre-application time prior to drug application. (B) Res-mediated inhibition of IGABA was almost saturated after 30 s of pre-application. The resting membrane potential of the oocytes was approximately -35 mV, and the oocytes were voltage-clamped at a holding potential of -80 mV. Each point represents the mean±S.E.M. (n=9~12/group).
|
Current-voltage relationship and voltage-independent inhibition of IGABA in oocytes that express GABAC receptors mediated by resveratrol
As shown in Fig. 4, the current-voltage relationship induced by GABA with voltage steps from -100 to +40 mV showed a slight rectification at positive potentials in oocytes with GABAC receptors. The reversal potential of GABAC receptors was Vr=-25.3±1.8 mV. (mean±S.E.M., n=6 from 3 frogs). Pre-treatment with resveratrol and GABA did not modify the potential reversal of GABAC receptor with a reduction of IGABA (n=6 from 3 frogs). The inhibitory effect of resveratrol on IGABA in oocytes that express GABAC receptors was independent of the membrane holding potential (Fig. 4B). Thus, resveratrol inhibited IGABA by 75.0±5.1, 79.6±3.1, 76.3±2.7, and 79.1±4.2% at -120, -90, -60, and -30 mV membrane holding potential in oocytes with GABAC receptor, respectively (Fig. 4B; n=9~12, from 3 frogs).
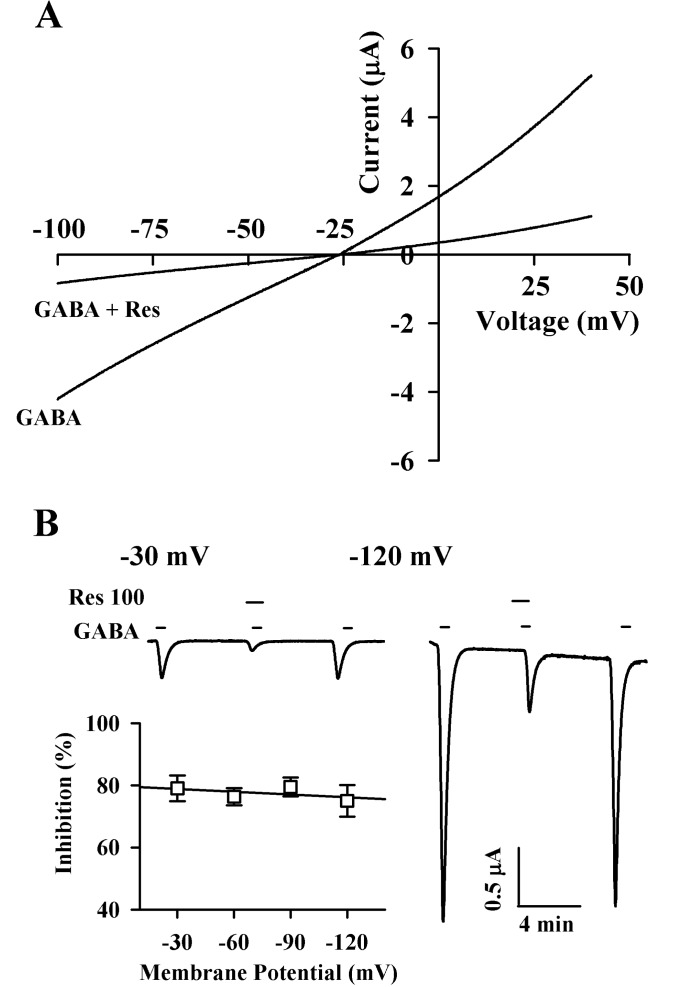 | Fig. 4Current-voltage relationship and voltage-independent inhibition by Res. (A) Current-voltage relationships of IGABA inhibition by Res in GABAC receptor-expressing oocytes. Representative current-voltage relationships were obtained using voltage ramps of -100 to +40 mV for 300 ms at a holding potential of -80 mV. Voltage steps were applied before and after application of 2 µM GABA in the absence or presence of 100 µM Res. (B) Voltage-independent inhibition of IGABA in the GABAC receptors by Res. Inset; the values were obtained from the receptors in the presence or absence of 100 µM Res at the indicated membrane holding potentials.
|
Noncompetitive inhibition of GABAC receptors by resveratrol
To further study the mechanism by which resveratrol inhibits IGABA in oocytes with GABAC receptors, we analyzed the effect of 100 µM resveratrol on IGABA evoked by various GABA concentrations on oocytes with GABAC receptors (Fig. 5). Pre-application of 30 or 100 µM resveratrol with different concentrations of GABA did not shift the dose-response curve of GABA to the right (ED50, from 2.2±0.3 to 2.4±0.1 and 3.1±0.2 µM and Hill coefficient, from 1.76 to 1.87 and 1.55) in oocytes expressing GABAC receptors, indicating that resveratrol regulates GABAC receptor channel activity in a non-competitive manner (n=9~12 from 3 frogs; Fig. 5).
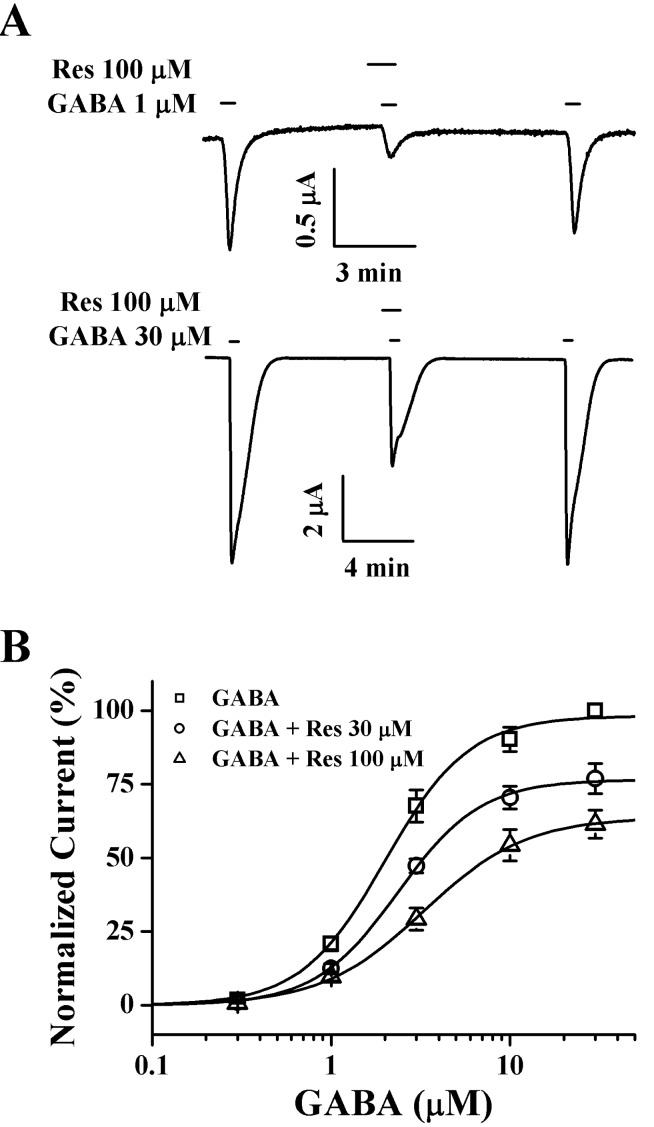 | Fig. 5Concentration-dependent effects of GABA on Res-mediated inhibition of IGABA. (A) The representative traces were obtained from the GABAC receptor-expressing oocytes. IGABA expression shown in the upper and lower panels were elicited at a holding potential of -80 mV by GABA at concentrations of 1 µM and 30 µM GABA respectively. (B) Concentration-response relationship of GABA with GABAC receptors treated with GABA (0.3~30 µM) alone or with GABA plus pre-application of 30 µM or 100 µM Res. The IGABA of oocytes expressing the GABAC receptors was measured using the indicated concentration of GABA in the absence (□) or presence of 30 µM (○) or 100 µM (▵) Res. Oocytes were exposed to GABA alone or to GABA with Res. Oocytes were voltage-clamped at a holding potential of -80 mV. Each point represents mean±S.E.M. (n=9~12/group).
|
Go to :
DISCUSSION
In the present study, we demonstrated that (1) co- or pre-treatment with resveratrol and GABA inhibited IGABA expression in human GABAC receptor-expressing oocytes in a reversible and concentration-dependent manner. (2) IGABA inhibition caused by resveratrol occurred in a non-competitive and voltage-independent manner in GABAC receptor-expressing oocytes, indicating that resveratrol could be associated with the inhibitory regulator of IGABA in GABAC receptor-expressing oocytes (Fig. 4 and 5).
However, our data was insufficient for elucidating the mechanisms for resveratrol inhibition of IGABA in GABAC receptor-expressing oocytes. The possibility that resveratrol may act as an open channel blocker of GABAC receptors seems unlikely because the inhibitory effect of resveratrol on IGABA in oocytes expressing GABAC receptors was not voltage-dependent (Fig. 4). It is known that open channel blockers such as local anesthetics or hexamethonium are strongly voltage dependent, due to the charge that they carry in the transmembrane electrical field [22-24].
Another possibility may be that resveratrol is a competitive inhibitor of GABAC receptors and inhibits the receptors by interacting with the receptor-binding site(s). Competition experiment data showed that the presence of resveratrol did not change the concentration of GABA in oocytes that express GABAC receptors without changing the Hill coefficient (Fig. 5). Thus, the non-competitive modulation of GABAC receptor channel activity of resveratrol shows that resveratrol might have other binding site(s) or interaction site(s), such as those of a non-competitive inhibitor, on the GABAC receptors.
Finally, the third possibility may be that resveratrol has binding sites that enable the regulation of GABAC receptor. In previous reports, we have demonstrated that the regulatory effects of resveratrol on homomeric 5-HT3 receptor channel activities were attenuated or abolished by site-directed mutations of amino acid residues of pre-transmembrane domain of 5-HT3 receptor [19]. On the basis of data from this study and that of the previous reports, we speculate that resveratrol achieves its effect through direct interactions with GABAC receptors. Further studies are required to identify resveratrol binding site(s) on the GABAC receptors.
Previous studies have shown that the effects of resveratrol on nervous system might be mediated by ligand-gated ion channels. For example, resveratrol-mediated neuroprotection against brain ischemia is inhibited by N-methyl-D-aspartate (NMDA) receptor antagonist [25]. Resveratrol also attenuates kainite-induced epilepsy [26]. In addition, resveratrol suppresses catecholamine secretion by inhibiting the α3β4 nicotinic acetylcholine receptor in adrenal medullary cells [27]. In addition, resveratrol potentiates 5-HT3A receptor channel activity [19]. Thus, although resveratrol is known as a neuroprotective agent, the mechanisms of resveratrol-mediated regulation of receptor or ion channel activities at the cellular level are poorly understood.
GABAC receptor is expressed in retina, thalamus, hippocampus, pituitary gland, and gut [7-10,28-30]. Its role may include visual processing, regulation of sleep-waking rhythms, pain perception, memory, learning, regulation of hormones, and neuroendocrine gastrointestinal secretion. Thus, although GABAC receptor channel activity might be closely related with the regulation of visual processing and other brain functions, we currently do not understand the inhibition of GABAC-receptor-mediated ion currents by resveratrol and its association with GABAC receptor-related functions in nervous system. Further studies are required to determine how in vitro resveratrol-mediated inhibition of IGABA is linked to GABAC receptor-related in vivo pharmacology in the nervous system.
In summary, we found that resveratrol, an active ingredient found in grapes, inhibits the GABAC receptor-mediated ion currents by interacting with sites that are distinct from the GABA-binding site(s), and that these results further indicate that the resveratrol-mediated GABAC receptor regulation might be the cellular basis for its effects on the nervous system.
Go to :
 XML Download
XML Download