

 PDF
PDF ePub
ePub Citation
Citation Print
Print


ABBREVIATIONS
ρCMB
ρ-chloromercuribenzoic acid
DEDA
dimethyl-eicosadienoic acid
EDG
endothelial differentiation gene
ERK
extracellular signal-regulated protein kinases
ESMCs
esophageal smooth muscle cells
JNK
c-Jun NH2-terminal kinases
LPA
lysophosphatidic acid
MAPK
mitogen-activated protein kinase
PKC
protein kinase C
PLC
phospholipase C
PLD
phospholipase D
PLA2
phospholipase A2
PTX
pertussis toxin
ROCK
Rho-associated kinase
SPC
sphingosinephosphorylcholine
SDS
sodium dodecyl sulfate
INTRODUCTION
Lysophosphatidic acid (LPA), sphigosine 1-phosphate (S1P), lysophosphatidylcholine (LPC) and sphingosylphosphorylcholine (SPC) are well-known lyso-type lipids. They are metabolic products of sphingomyeline (N-acylsphingosine-1-phosphocholine) which is a component of cell membrane and lipoprotein. Till now many research about the lyso-type lipids has been reported [1]. Among the lyso-type lipids, LPA has recently emerged as an intracellular phospholipids messenger. LPA common name is monoacyl-snglycero-3-phosphate. Diverse cellular actions of LPA on many cell types include platelet aggregation, tumor cell invasion, wound healing, vascular remodeling, neurite retraction, inhibition/reversal of differentiation, membrane depolarization and release of neurotransmitters, regulation of cell proliferation, protection from apoptosis, modulation of chemotaxis and transcellular migration [2-4]. Moreover, LPA has been recently evaluated by its clinical significance in esophagus. LPA-specific phosphatase is used as prognostic factor for patients with esophageal squamous cell carcinoma [5]. LPA has been also known to modulate contraction of various types of smooth muscle including airway and uterine smooth muscle [6-9]. However, little is known about the signal transduction mechanism of the LPA-induced contraction in gastrointestinal tract.
Because LPA does not penetrate cells, most of the effects of LPA are now thought to be receptor-mediated [1,3,10-12]. LPA, S1P, and SPC activate each specific members of the G protein-coupled receptor (GPCR) superfamily [13]. Based on sequence homology, the mammalian LPA receptors belong to the so-called endothelial differentiation gene (EDG) subfamily of the GPCR superfamily of seven-transmembrane domain receptors. LPA has been known to interact with at least three GPCRs in mammals, such as LPA1/Edg-2, LPA2/Edg-4, and LPA3/Edg-7 [14-17]. In common with most GPCRs, LPA receptors undergo rapid ligand-induced internalization from the plasma membrane. It was not confirmed that these kinds of LPA receptors exist in feline esophageal smooth muscle cells (ESMCs).
LPA-mediated responses attributed to an intracellular second messenger action. Pertussis toxin (PTX) pretreatment which specifically inactivates Gi/o-type G protein, inhibits many responses to LPA but, it is not specific for a particular receptor subtype. LPA receptor was found to stimulate a proliferation through PTX-sensitive G proteins (Gi/o) and a cytoskeletal remodeling through PTX-insensitive ones (G12/13) and Rho. In addition, LPA receptor coupled with PLC activation possibly through a PTX-insensitive Gq/11 [18]. The previous study in our lab demonstrated the presence of immunoreactive bands of 150 kDa with PLCβ1 and PLCβ3 antibodies, and a 145 kDa band with PLCγ1 antibody in dispersed esophageal smooth muscle cells [19].
Protein kinase C (PKC) plays a pivotal role in cell signaling by relaying information from lipid mediators to protein substrates [20,21]. In esophageal smooth muscles, the activation of PKC is linked to mitogen-activated protein kinases (MAPKs) pathway which leads to the muscle contraction [22]. PKC is present in the cell cytoplasm and upon stimulation, it translocates to the membrane fraction or particulate [23]. It can be hypothesized that LPA receptor and PKC act on a target upstream of activated MAPK-induced smooth muscle cell contraction.
Several GPCRs, including LPA receptors, have been determined to mediate the signaling pathway to activate G-protein. Small GTP-binding protein (G-protein) including Ras, Rho, Rac and their downstream MAPKs play a central role in cellular responses [24]. The best-characterized members of the MAPK superfamily of protein kinases are the ERK1/2. ERK1/2 can be activated by many different stimuli and are involved in smooth muscle cell contraction [25].
RhoA/Rho kinases (ROCK) are critical for the Ca2+ sensitization of 20-kDa myosin regulatory light chain (MLC20) in smooth muscle cells. ROCK, as one of the downstream effectors of Rho GTPases, plays an important role in smooth muscle contraction. Various vasoactive factors stimulate RhoA and ROCK, leading to enhanced vasoconstriction and migration of vascular smooth muscle cells [26]. In the present study, we therefore investigated the involvement of specific LPA receptors in LPA-induced contraction of cat ESMCs, and examined the signaling pathways initiated by the receptor to assess the effect of LPA as a ligand.
Go to : 
METHODS
Materials
DEDA, PD98059, SB202190, EDG-7 receptor antibodies were purchased from Calbiochem (San Diego, CA); EDG-2 receptor antibodies was purchased from Abcam (cambridge, UK); PKC isozyme antibodies (-βII, -γ, and -ε) from Santa Cruz Biotechnology (Santa Cruz, CA); GF109203X from Tocris (Ellisville, MO, USA); C3 exoenzyme was purchased from Merck (Darm stadt, Germany). Y27632 was purchased from Biomol research laboratories (PA, US). Rainbow prestained molecular weight marker from Amersham (Arlington Heights, IL, USA); enhanced chemiluminescence agents from PerkinElmer Life Sciences (Boston, MA); nitrocellulose membrane from BioRad (Richmond, CA, USA); sodium dodecyl sulfate (SDS) sample buffer from Owl scientific, Inc (Woburn, MA); phosphate-buffered saline (PBS) from Boehringer Mannheim (Indianapolis, IN, USA). LPA, kreb buffer, Horseradish peroxidase-conjugated goat anti-rabbit antibody, 4-(2-hydroxyethyl)-1-piperazine-N'-2-ethane sulfonic acid (HEPES), collagenase type F, ammonium persulfate, ponseu S, bovine serum albumin (BSA), pertussis toxin (PTX), U73122, ρCMB, ethylene glycolzbis-(β-aminoethyl ether)-N,N,N',N'-tetraacetic acid (EGTA), ethylenediamine tetraacetic acid (EDTA), and other reagents were purchased from Sigma (St Louis, MO).
Preparation of feline esophageal smooth muscle squares
All animal experiments were approved by the Institutional Animal Care and Use Committee of Chung-Ang University, in accordance with the guide for the Care and Use of Laboratory Animals in Seoul, Korea. Adult cats of either sex weighing between 2.5 and 4 kg were used in this study. The cats were anesthetized with 12.5 mg/0.25 ml/kg Zoletil 50 and then abdomen was opened with a midline incision. The esophagus was excised and cleaned and then maintained in Krebs buffer of following composition (mM): NaCl 116.6, NaHCO3 21.9, NaH2PO4 1.2, KCl 3.4, CaCl2 2.5, glucose 5.4 and MgCl2 1.2. The esophagus was opened along the lesser curvature. The location of the squamocolumnar junction was identified and the mucosa was peeled off. Then, the submucosal connective tissue on the smooth muscle was removed by micro spring scissor. The cleaned smooth muscle was cut into less than 0.5 mm thick slices with a Stadie Riggs tissue slicer (Thomas Scientific Apparatus, Philadelphia, PA, USA). The slices were cut into 2×2 mm tissue squares by scissors.
Preparation of dispersed smooth muscle cells
Single muscle cells were isolated as previously described [27,28]. Muscle strips were incubated overnight in normal potassium-HEPES buffer containing 1 mg/ml papain, 1 mM dithiothreitol, 1 mg/ml bovine serum albumin (BSA) and 0.5 mg/ml collagenase (type F, Sigma) and equilibrated with 95% O2 - 5% CO2 to maintain pH 7.0 at 31℃. The composition of the normal potassium-HEPES buffer was 1 mM CaCl2, 250µM EDTA, 10 mM glucose, 10 mM HEPES, 4 mM KCl, 131 mM NaCl, 1 mM MgCl2 and 10 mM taurine. Next day we warmed up the tissue at room temperature for 30 min and incubated the tissue in a water bath at 31℃ for 30 min. After incubation, the digested tissue was poured out over a 360-µm Nitex filter, rinsed in collagenase-free HEPES buffer to remove any trace of collagenase, and then incubated in this solution at 31℃, gassed with 95% O2 - 5% CO2. The cells were allowed to dissociate freely for 10 to 20 min. Suspensions of single muscle cells were harvested by filtration through 500-µm Nitex mesh [27]. Before beginning the experiment, the cells were kept at 31℃ for at least 10 min to relax the cells. Throughout the entire procedure, care was taken not to agitate the fluid to avoid cell contraction in response to mechanical stress.
Preparation of permeabilized smooth muscle cells
Smooth muscle cells were permeabilized to diffuse the agents such as C3 exoenzyme, Y27632 or PKC antibody, which do not diffuse across the intact cell membrane. After completion of the enzymatic phase of the digestion process, the partly digested muscle tissue was washed with an enzyme-free cytosolic buffer of the following composition: 20 mM NaCl, 100 mM KCl, 5.0 mM MgSO4; 0.96 mM NaH2PO4; 1.0 mM EGTA and 0.48 mM CaCl2 and 2% bovine serum albumin. The cytosolic buffer was equilibrated with 95% O2 - 5% CO2 to maintain a pH of 7.2 at 31℃. Muscle cells dispersed spontaneously in this medium. The cytosolic buffer contained 0.48 mM CaCl2 and 1 mM EGTA, yielding 0.18µM free Ca2+ as calculated according to [29]. After dispersion, the cells were permeabilized by incubation for 3 min in cytosolic buffer that contained saponin (75µg/ml). After exposure to saponin, the cell suspension was spun at 350 g, and the resulting pellet was washed with saponin-free modified cytosolic buffer that contained antimycin A (10µM), ATP (1.5 mM) and an ATP-regenerating system that consisted of creatine phosphate (5 mM) and creatine phosphokinase (10 units/ml). After the cells were washed free of saponin, they were resuspended in modified cytosolic buffer.
Measurement of contraction by scanning micrometry
Contraction of isolated muscle cells was measured by scanning micrometry [31]. An aliquot of cell suspension containing 104 cells/ml was added to HEPES medium containing the test agents. The reaction stopped by accustain treatment. Length of individual cells was measured before and after the treatment of contractile agents by scanning micrometry, phase contrast microscope (model ULWCD 0.30 Olympus, Tokyo, Japan) and digital closed-circuit video camera (CCD color camera, Toshiba, Tokyo, Japan) connected to a Macintosh computer (Apple, Cupertino, CA) with a software program, NIH Image 1.57 (National Institutes of Health, Bethesda, MD). The average length of cells encountered randomly in successive microscopic fields. The experiments were repeated in at least three animals. Contraction was expressed as the percentage decrease in mean cell length compared to the control.
Polyacrylamide gel electrophoresis and immunoblotting
The identification of EDG receptors in esophageal smooth muscle was performed by Western blot analysis. Intact smooth muscle cells were homogenized in a lysis buffer containing 20 mM Tris, 0.5 mM EDTA, 0.5 mM EGTA, 10µg/ml leupeptin, 10µg/ml aprotinin, 10 mM β-mercaptoethanol (pH 7.5). Sample homogenates were then centrifuged for 10 min at 4℃, and the supernatants were collected. Aliquots were subjected to electrophoresis on a 10% SDS-polyacrylamide gel and transferred onto a nitrocellulose membrane. Transfer of proteins to the nitrocellulose membrane was confirmed with Ponseau S staining reagent. To block nonspecific binding, the nitrocellulose membrane was incubated in 5% nonfat dry milk in phosphate-buffered saline (PBS) for 60 min followed by three rinses in milk-free buffer. Incubation with 1:1,000 dilution of antibody raised against each EDG receptor was done for 1 h with shaking followed by three washes with antibody-free buffer. This was followed by 60-min incubation in horseradish peroxidase-conjugated goat anti-rabbit antibody (dilution 1:2,000). Detection was achieved with an enhanced chemiluminescence agent. Molecular mass was estimated by comparison of sample bands with a prestained molecular mass marker.
Protein assay
For equal amounts of the protein from each sample were resolved on a SDS-polyacrylamide gel by electrophoresis, the protein concentration of supernatant was determined by the Bradford reagents according to the instructions of the manufacturer (Bio-Rad Chemical Division, Richmond, California). The absorbance was measured spectrophotometrically at a wavelength of 595 nm.
Statistical analysis
Data are expressed as means±SEM of separate experiments and the statistical differences between means were determined by Student's t-test.
Go to :
RESULTS
Preparation of dispersed esophageal smooth muscle cells
To check the dispersing process of esophageal smooth muscle in collagenase buffer, the dissected smooth muscle strip in collagenase buffer was observed in every 5minutue by microscopy during incubation. It was observed that esophageal smooth muscle cells (ESMCs) were being separated from esophageal smooth muscle tissue squares. Freshly isolated esophageal smooth muscle cells were presented in spindle shape with diverse length range of 38~113µm.
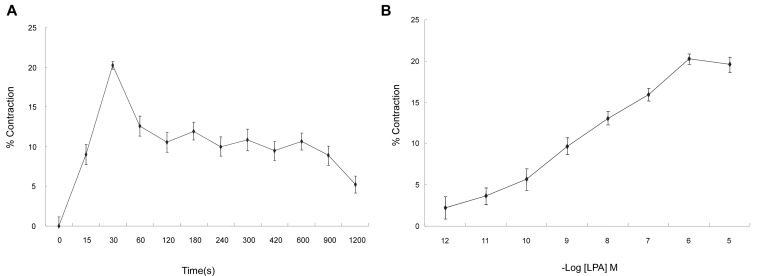
LPA-induced contraction of isolated ESMCs
Freshly isolated esophageal smooth muscle cells were stimulated with 10-6 M LPA for up to 1,200 s. LPA-induced contraction of smooth muscle cells that was peaked at 30 s and them slowly declined (Fig. 1A). The response to LPA was concentration-dependent (Fig. 1B). The cells were stimulated with 10-12 to 10-5 M LPA for 30 s. Maximal contraction occurred at 10-6 M. On the basis of these data, ESMCs were exposed to 10-6 M LPA for 30 s in most of later experiments.
Detection of LPA receptors in ESMCs and the effects of Ki16425 and PTX on contraction induced by LPA
The previous study determined S1P receptor subtypes that belong to EDG receptor subfamily like LPA receptors by western blot analysis [32]. To identify the LPA receptor subtypes in ESMCs, western blot analysis was performed. Western blot analysis of homogenates obtained from dispersed smooth muscle cells using antibodies to LPA receptor subtypes demonstrated the presence of immunoreactive protein bands corresponding to LPA1 (EDG2) and LPA3 (EDG7) antibody at 50 kDa and 44 kDa (Fig. 2A).
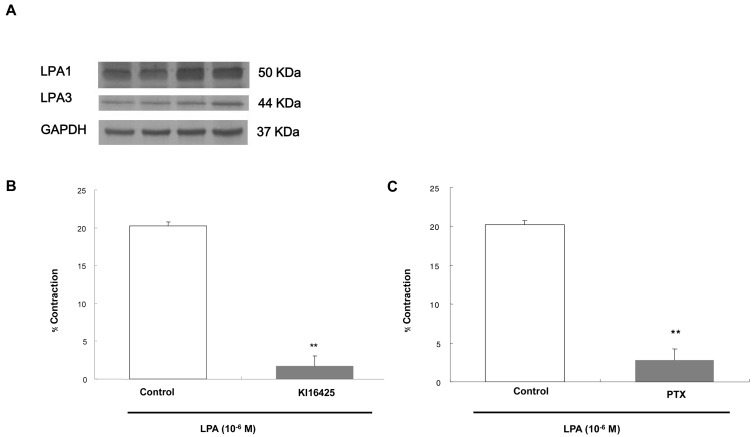 | Fig. 2(A) Identification of LPA receptor subtype in esophageal smooth muscle cell by western blot analysis. LPA1 and LPA3 receptors were detected at 50 kDa and 44 kDa. The blot also was probed with anti-GAPDH antibody as a loading control. (B) Effect of receptor antagonist on the contraction induced by LPA (10-6 M). The intact cells were preincubated with KI16425 (10-6 M), LPA receptor 1/3 antagonist, for 10 min. (C) Effect of PTX on the contraction induced by LPA (10-6 M). The intact cells were preincubated with PTX (400 ng/ml) for 60 min. Data are expressed as means±SEM of three independent experiments. **p<0.01 vs. control, two-tailed t-test.
|
To estimate the contractile signal mediated by LPA1 and LPA3 receptor, the intact cells were preincubated for 10min with Ki16425 (10-6), selective LPA1/3 receptor antagonist. LPA-induced contraction was almost completely blocked by Ki16425. Fig. 2B shows that LPA-induced contraction of ESMCs was significantly inhibited by Ki16425, from 20.24±0.51 to 1.70±1.38.
It has been known that LPA has its own receptor coupled with PTX-sensitive G-protein. To test the effect of PTX on contraction by LPA, the cells were preincubated with PTX (400 ng/ml) for 60 min. Fig. 2C shows that LPA-induced contraction of esophageal smooth muscle cells were significantly abolished by PTX, from 20.24±0.51 to 2.79±1.45. This implies that LPA-induced contraction in ESMCs is coupled to a PTX-sensitive G protein.
LPA-induced contraction is mediated by PLC
LPA-induced contraction of ESMCs was not affected by PLA2 inhibitor DEDA (10-5 M) and PLD inhibitor ρCMB (10-5 M), but significantly abolished by PLC inhibitor Neomycin and U73122 (10-6 M). Percent decrease in cell length was as follows: 20.24±0.51 vs. 7.30±1.23, 8.61±1.08, 18.04±1.26 or 19.04±1.17 in the cells preincubated with Neomycin, U73122, ρCMB or DEDA, respectively (Fig. 3). These results suggest that contraction of ESMCs is partially mediated by phosphatidylinositol-specific PLC.
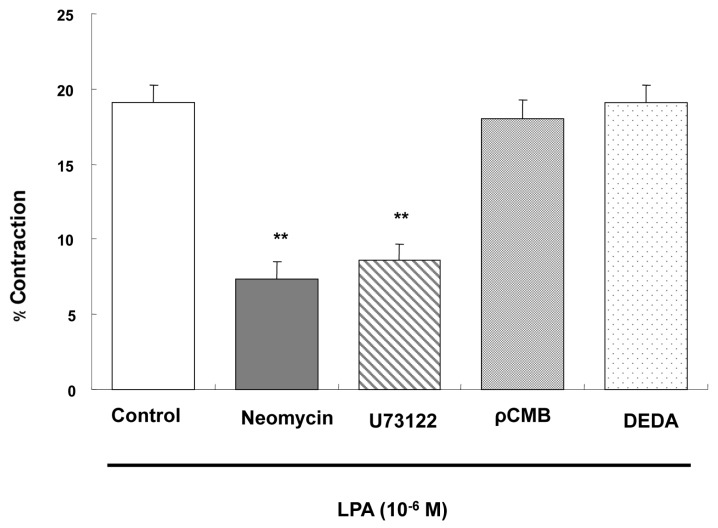 | Fig. 3Effect of phospholipases on the contraction induced by LPA (10-6 M). The intact cells were preincubated with PLC inhibitors neomycin (10-6 M) or U73122 (10-6 M) for 10 min, PLD inhibitor ρCMB (10-5 M) for 10 min, or PLA2 inhibitor DEDA (10-5 M) for 1 min. Data are expressed as means±SEM of three independent experiments. **p<0.01 vs. control, two-tailed t-test.
|
Determination of PKC isozyme involvement in LPA-induced contraction
Cells were preincubated with PKC inhibitor GF109203X (3×10-6 M) for 10 min before the addition of LPA. LPA-induced contraction of ESMCs was inhibited by PKC inhibitor, from 20.24±0.51 to 6.48±1.43 (Fig. 4A). These results suggest that LPA-induced esophageal smooth muscle contraction is mediated by the activation of PKC cascades.
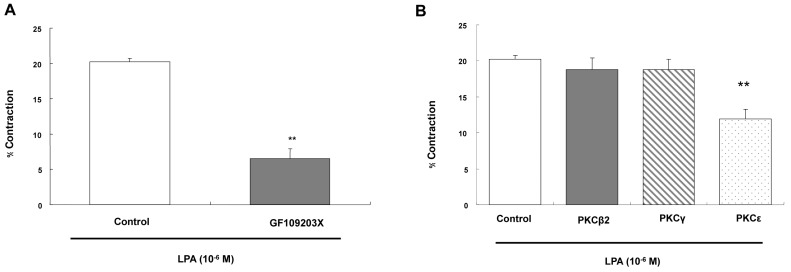 | Fig. 4(A) Effect of protein kinase C inhibitor on the contraction induced by LPA (10-6 M). The intact cells were preincubated with PKC inhibitor GF109203X (3×10-6 M) for 10 min. (B) The permeabilized cells were preincubated with antibodies raised against PKC isozymes for 1 h. PKCε antibody reduced the contraction. Data are expressed as means±SEM of three independent experiments. **p<0.01 vs. control, two-tailed t-test.
|
In our previous study, western blot analysis showed that PKC isozymes, including the PKC-βII, -γ, and -ε isozymes, are present in the smooth muscle of the esophagus [33]. To test whether the PKC-mediated contraction is isozyme-specific, the permeabilized cells were used to examine the effect of PKC isozyme antibodies on contraction induced by LPA. Figure 4B shows that LPA-induced contraction of permeabilized esophageal smooth muscle cells was significantly inhibited by antibodies raised against PKC-ε (1±200), from 20.24±0.51 to 11.94±1.37, but not by antibodies raised against the PKC-βII or -γ isozyme (18.79±1.61 or 18.76±1.44).
Involvements of PKC and MAPK on LPA-induced contraction in ESMCs
To determine whether MAPK is required for LPA-induced contraction, specific MAPK inhibitors were used. Preincubation of cells with ERK1/2 inhibitor PD98059 for 30 min blocked the contraction induced by LPA. The selective inhibitor of JNK, SP600125, inhibited the contraction too. However, preincubation of p38 MAPK inhibitor SB202190 for 30 min did not inhibit LPA-induced contraction. Percent decrease in cell length was as follows: 20.24±0.51 vs. 9.30±1.27, 11.60±1.15 or 16.78±1.00 in the cells preincubated with PD98059, SP600125 or SB202190, respectively (Fig. 5A). The data suggests that LPA-induced contraction is mediated via the ERK1/2 or JNK pathway.
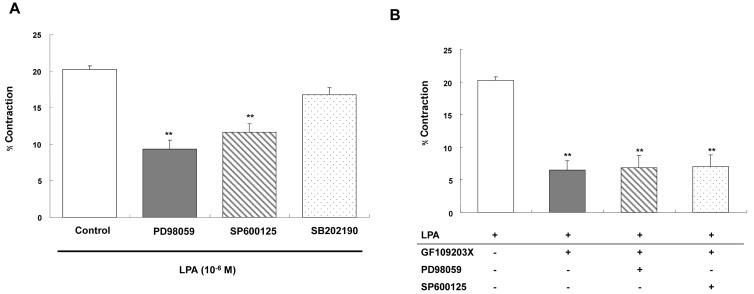 | Fig. 5(A) Role of MAPK on LPA-induced smooth muscle cells contraction. The intact cells were pretreated with ERK1/2 inhibitor PD98059 (10-5 M), JNK inhibitor SP600125 (10-5 M), or p38 MAPK inhibitor SB202190 (10-5 M) for 30 min. (B) Synergistic effect of PKC and MAPK on LPA-induced contraction. The PKC inhibitor was co-treated with ERK1/2 or JNK inhibitor. Data are expressed as means±SEM of three independent experiments. **p<0.01 vs. control, two-tailed t-test.
|
To test the synergic effect of PKC and MAPK on LPA-induced contraction in ESMCs, the PKC inhibitor was co-treated with ERK1/2 or JNK inhibitor. Percent decrease in cell length was as follows: 20.24±0.51 vs. 6.5±1.43, 6.83±1.88 or 7.03±1.76 in the cells preincubated with GF109203X only, GF109203X with PD98059, or GF109203X with SP600125, respectively (Fig. 5B). Therefore, the cotreatment of cells did not show synergistic effects, suggesting that these kinases are involved in the same signaling pathway.
Involvement of Rho on LPA-induced esophageal smooth muscle cell contraction
A recent study found that RhoA/Rho-kinase are involved in vascular smooth muscle cell contraction [34]. Therefore, we investigated the involvement of RhoA in the LPA-induced contraction of ESMCs. RhoA inhibitor C3 exoenzyme and ROCK inhibitor Y27632 were tested. The permeabilized cells were preincubated with C3 exoenzyme (10µg/ml) for 30 min, or with Y27632 (10-5) for 10 min. LPA-induced contraction of permeabilized ESMCs was reduced by C3 exoenzyme and by Y27632, from 20.24±0.51 to 10.02±1.44 and from 20.24±0.51 to 10.97±1.13 (Fig. 6). Thus, the RhoA inhibitor and the ROCK inhibitor significantly, but not completely, reduced the contraction.
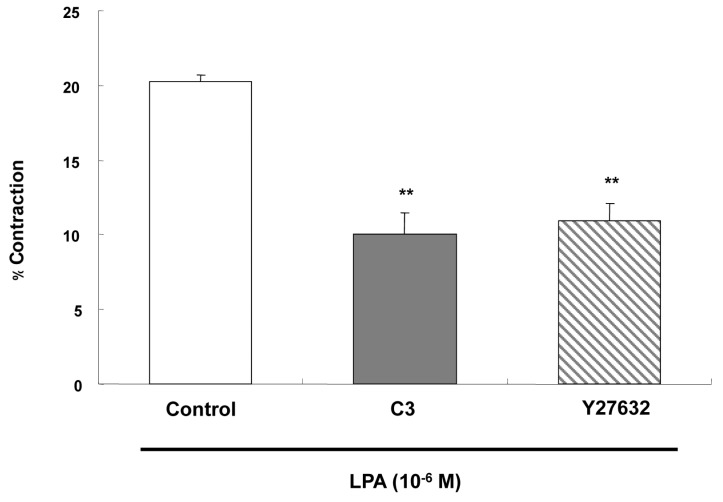 | Fig. 6Effects of RhoA inhibitor C3 exoenzyme and ROCK inhibitor Y27632 effect on the contraction induced by LPA (10-6 M). The permeabilized smooth muscle cells were preincubated with C3 exoenzyme for 30 min (10µg/ml), or Y27632 (10-5 M) for 10 min. Data are expressed as means±SEM of three independent experiments. **p<0.01 vs. control, two-tailed t-test.
|
Go to :
DISCUSSION
In the present study, we investigated the signaling pathway on lysophosphatidic acid (LPA)-induced contraction in cat esophageal smooth muscle cells. LPA induced a dose-dependent manner contractile effect. We found that the LPA-induced contraction depends on activation of PTX-sensitive G protein and PLC via LPA receptors (1/3), leading to stimulation of a PKC-ε pathway, which subsequently activates ERK and JNK.
In this study, the contractile reaction was stopped by accustain. In our previous study, we had used formalin or acrolein, instead of accustain [35]. The accustain was less cytotoxic than formalin and acrolein when preliminary test was done. Besides, accustain was not different from formalin in fixing ability for 24 h and did not affect the contraction (Data not shown).
The three LPA receptors of the EDG family show about 50% sequence similarity to one another, with their C-terminal tails being most divergent [1]. Candidate high-affinity LPA receptors were initially detected by photo-affinity labeling experiments [36]. LPA1 receptor is the most widely expressed and best characterized subtype, whereas LPA2 receptor and LPA3 receptor have a somewhat more restricted distribution pattern [37]. LPA1 gene is expressed in many tissues, including testes, lung, heart, intestine, and stomach in adult mouse and human. LPA2 gene is expressed in many tissues too. However, little or no expression was detectable in small intestine or colon in mouse and human. Mouse LPA3, like LPA2, is expressed in testes, kidney, lung, small intestine and low level in stomach. But human LPA3 expression was not detected in small intestine and colon [1,13]. For those reasons, the detection of only LPA1 and LPA3 receptors was performed by western blot analysis in this study. In addition, LPA receptor-selective antagonists, which have recently become commercially available, are important tools for identifying the subtype of LPA receptors. Ki16425, which was used in this study, is a LPA receptor antagonist that shows a preference for LPA1/3 over LPA2 [38].
In gastrointestinal tracts, activated phospholipase produce second messengers, such as arachidonic acid, inositol-1,4,5-triphosphate (IP3) and diacylglycerol (DAG), by degrading phospholipids to induce contraction. LPA receptors are coupled with PLC activation possibly through a PTX-insensitive Gq/11 [18]. Our previous paper shows that western blot analysis of homogenates obtained from dispersed smooth muscle cells using polyclonal antibodies to PLC isozymes demonstrated the presence of immunoreactive protein bands corresponding to 150 kDa (PLCβ1 and PLCβ3 antibodies), and a 145 kDa (PLCγ1) [19]. Besides, the contraction induced by S1P, one of lysolipids, like LPA was inhibited by PLC β3 antibody [32]. The LPA-induced contraction significantly abolished by PLC inhibitors. These results suggest that LPA-induced contraction of ESMCs be partially mediated by phosphatidylinositol-specific PLC.
PKC is a family of homologous serine and threonine protein kinases. PKC is present in the cell cytoplasm, and upon stimulation, it translocates to the membrane fraction or particulate [23]. PKC plays an important role in cell signaling by relaying information from lipid mediators to protein substrates [21]. Direct introduction of active PKC causes contraction of smooth muscle cell [30,39]. Recent studies have demonstrated that PKC α translocates from the cytoplasm to membrane on stimulation by contractile agonist during smooth muscle cell contraction, and the agonist-stimulated PKC translocation was observed by western blot or immunofluorescence labeling [40]. The present study also suggests that PKC pathway is involved in the LPA-induced contraction, and the PKC-mediated contraction is isozyme-specific.
PKC isozyme can be divied into three groups, depending on their calcium and phospholipids requirements for activation; classical or conventional PKCs (cPKC), including α, β1, β2, γ which are calcium- and phospholipids dependent; new PKCs (nPKC), including δ, ε, η, θ and µ, which are calcium-independent and phospholipids-dependent; and atypical PKC (aPKC), including ζ and λ, which are calcium- and phospholipid-independent [41]. The present study suggests that PKC ε play a role in the LPA-induced ESMC contraction.
It has been known that MAPK mediates LPA-induced cell responses [42]. There are 14 MAPKs in mammalian cells that can be divided into four groups: classical MAPKs (extracellullar signal regulated kinase (ERK)1 and ERK2), c-Jun NH2-terminal kinases (JNK1, JNK2, JNK3), and p38 MAPK (p38α, p38β, p38γ and p38δ) and atypical MAPKs (ERK3, ERK5 and ERK8) [26]. The compounds of PD98059, SP600125, and SB202190 have been identified as highly selective, potent and cell permeable inhibitors of ERK1/2, JNK, and p38 MAP kinase, respectively [43,44]. SB202190 is an important tool for the study of p38 MAP kinase function both in vivo and in vitro [45]. In the present study, the LPA-induced contraction was blocked by ERK1/2 and JNK inhibitors, but not by p38 MAPK inhibitor. Since MAPK activation is often a part of downstream signaling of PKC or Rho activation, PKC inhibitors were cotreated with ERK1/2 or JNK inhibitor. The cotreatment did not show synergistic effects, suggesting that these kinases are involved in the same signaling pathway.
The data also suggest that RhoA/ROCK play a significant role for the maintenance of contractile state of the smooth muscle cell. Inactive RhoA in the cytoplasm remains as RhoA-GDP complexed with Rho guanine nucleotide dissociation inhibitor (GDI) [46]. Guanine nucleotide exchange factor catalyze the exchange of GDP-RhoA-GDI to active RhoA-GTP that associates with plasma membrane. RhoA-GTP binding to Rho binding domain of ROCK leads to autophosphorylation and activation of ROCK [47,48]. Activated ROCK inhibits myosin light chain phosphatase (MLCP). MLCP causes dephosphorylation of MLC20. MLCP is a heteromeric enzyme that present in the smooth muscle [30].
LPA increase the intracellular free calcium concentration [49]. The relationship of contractile responses by LPA and calcium in esophageal smooth muscle cells would be further investigated in the near future. In conclusion, the LPA-induced contraction in feline esophageal smooth muscle cells seems to be mediated by LPA receptor (1/3), coupled to PTX-sensitive G protein, resulting in the activation of PLC, PKC-ε pathway, which subsequently mediates the activation of ERK and JNK. The data also suggest that RhoA/ROCK is involved in the LPA-induced contraction (Fig. 7).
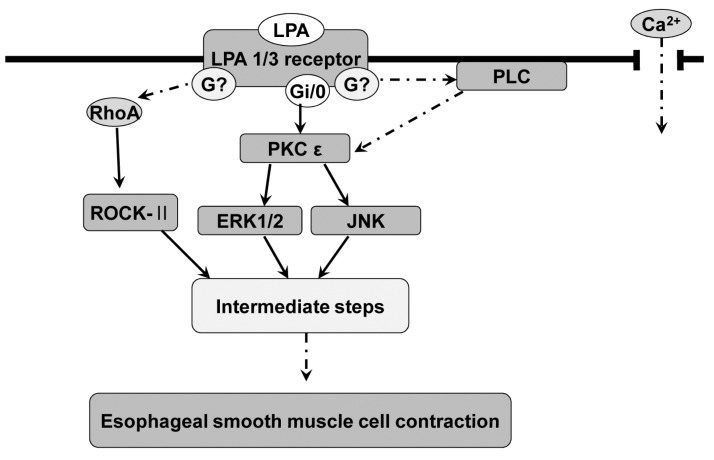 | Fig. 7Expected intracellular signal pathways of LPA-induced contraction in esophageal smooth muscle cell. LPA-induced contraction seems to be mediated by LPA receptor (1/3), coupled to PTX-sensitive G protein, resulting in the activation of PLC, PKC-ε pathway, which subsequently mediates the activation of ERK and JNK. The data also suggest that RhoA/ROCK is involved in the LPA-induced contraction.
|
Go to :
 XML Download
XML Download