

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Tooth pulpal pain (toothache) is one of the most common pains experienced by human and it is well known that nociceptive signals from tooth are conducted into the thalamic specific relay nuclei through trigeminal spinal subnuclei; subnucleus oralis, subnucleus interpolaris and subnucleus caudalis [1]. Besides, tooth pulpal pain affects the cardiovascular system, resulted in increasing both arterial blood pressure (BP) and heart rate (HR), potentially inducing stroke in hypertensive patients [2,3]. This cardiovascular reflex elicited in response to noxious stimulation of trigeminal nerves is mediated by trigeminal-autonomic reflex affected by other brain regions outside the spinal trigeminal nucleus since trigeminal nuclei do not appear to have intrinsic autonomic functions. However, the central mechanisms underlying cardiovascular reflex elicited by dental pulpal pain is little understood.
Ginseng, the root of panax ginseng C.A Meyer (Araliaceae), is a well-known traditional herbal medicine that has been shown to exhibit a variety of medicinal efficacies. The main molecular components responsible for the action of ginseng are ginsenosides which are also called a ginseng saponin. The ginsenosides have a four-ring, steroid-like structure with sugar moieties attached and show similar properties to various neurotransmitters. They also have an antinociceptive effect on various animal models [4-8]. In practice, ginseng has been used to alleviate a variety of pain in traditional folk medicines since administration of ginseng extracts promptly produces antinociception and prevents the opioid tolerance caused by chronic treatment with morphine at spinal cord [7]. Moreover, ginsenosides recently were known to inhibit voltage-dependent channels in peripheral sensory neurons [9,10]. These reports indicate that ginseng plays an important role in modulating pain perception in the central nervous system and peripheral nervous system. However, effects of ginsenosides on tooth pulpal pain and trigemino-cardiovascular reflex were not studied using c-Fos expression, a reliable marker to identify nociceptive neuron or specific neural pathways activated by noxious stimuli in animal model. This study investigated the effects of ginsenosides on tooth pulpal pain and trigemino-cardiovascular reflex elicited by noxious tooth pulpal stimulus for c-Fos in rats.
Go to : 
METHODS
Animal preparation and drug administration
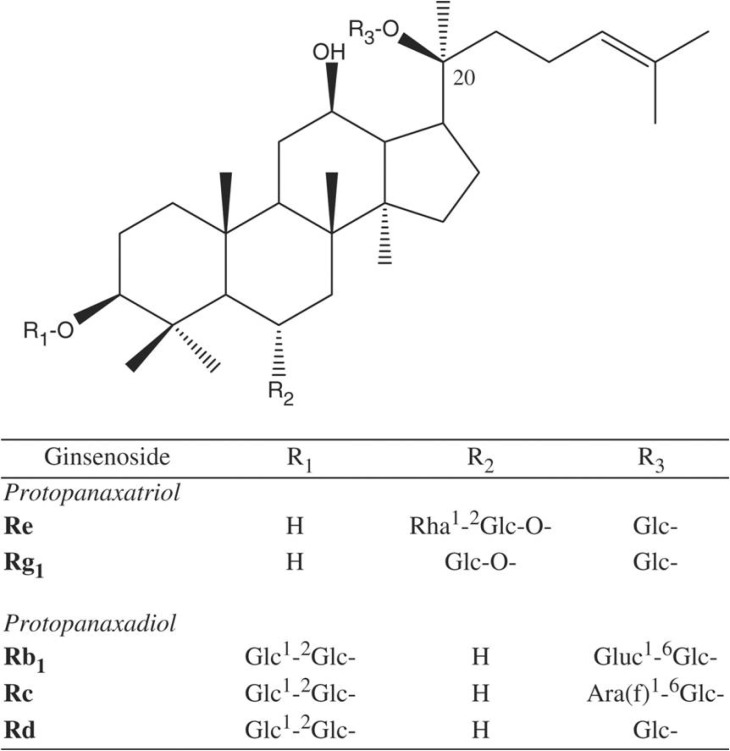
Male sprague Dawery rats (300~350 g) were anesthetized with sodium pentobarbital (100 mg/kg, i.p.). Both upper and lower incisors were cut by bone cutter, and noxious stimulation was given to the exposed pulp by intrapulpal injection of 2 M KCl. Naloxone, an opioid antagonist, was purchased from Sigma (St. Louis, Mo, USA) and ginsenosides are a gift from the Korea Ginseng and Tobacco Research Institute. Ginsenosides are a mixture of various types of individual ginsenosides, and the ratios of various ginsenosides are as follows; Rb1 (18.2%), Rb2 (9.07%), Rc (9.65%), Rd (8.24%), Re (9.28%), Rf (3.48%), Rg1 (6.42%), Rg2 (3.63%), Rg3 (4.70%), Ro (3.82%), Ra (2.91%) and other minor ginsenosides (Fig. 1). Ginsenosides were injected into peritonium 30 min before intrapulpal injection of 2 M KCl. Naloxone (5 mg/kg) was injected intraperitoneally 20 min before administration of ginsenosides. All procedures were performed in accordance with the ethical standards suggested by the animal care committee in Chonnam National University.
Immunohistochemistry for c-Fos
The method for analysis of c-Fos expression in this study was performed according to our previous study [11]. The rats were intracardially perfused with freshly prepared cold (4℃) 4% araformaldehyde in 0.1 mol/l phosphate buffer (pH 7.4) 1.5 h after NS. The brain was removed, fixed for 2 h, and cryoprotected in 18% sucrose for two days. It was then frozen on dry ice and sectioned with 30 µm thickness on a cryostat microtome.
Immunohistochemical staining was carried out using Vectastain Elite ABC kit (Vector Laboratories, Lexington, KY, USA). The tissue sections were incubated in 3% H2O2 for 30 min to block endogenous peroxidase and in 10% normal goat serum for 30 min to block non-specific reaction. After a short incubation with 2% Triton-X100, the sections were rinsed and incubated with the primary antibody to the rat c-Fos (Santa Cruz Biotechnology, Santa Cruz, CA, USA) (1:10,000) at 4℃ for 48 h. After rinsing with phosphate-buffered saline (PBS), they were incubated with a biotinylated secondary antibody (1:500 dilution) with 0.3% Triton-X100 in PBS for 1 h at room temperature. They were then rinsed with PBS and incubated with avidin-biotin-horseradish peroxidase complex solution (1:115 dilution with 0.3% Triton-X100 in PBS) at room temperature for 1 h. They were again rinsed with PBS and developed with 0.05% diamino benzidine acid as a chromogen. They were mounted on gelatin-coated slides, dried, dehydrated with absolute alcohol, and cover-slipped with Permount. The numbers of c-Fos immunoreactive neurons were counted with eye on the light microscope. We observed c-fos expression in same level of trigeminal spinal nucleus involving subnucleus codalis, interpolaris and oralis and calculated the mean number of c-Fos positive nurons.
Measurement of arterial blood pressure and heart rate
Male Sprague Dawley rats (300~350 g, n=12) were anesthetized with sodium pentobarbital (100 mg/kg, i.p.) immediately followed by 20% urethane (1.0 g/kg, i.p.). The femoral artery was cannulated and coupled to a statham physiological pressure transducer. Arterial pressure measured from the transducer was recorded on polygraph (Grass Co., USA) before and after noxious stimulation of the upper and lower incisors pulp. Heart rate (HR) was calculated from the peak to peak interval of the arterial blood pressure (BP) pulses. In some cases, a tracheostomy was performed to maintain airway. Body temperature was maintained at 37±5℃ on the heating pad.
Statistical analysis
Five animals were used in each group for quantification and results are presented as mean±SEM. Statistical analysis was performed using one-way analysis of variance (ANOVA) followed by Student's t test.
Go to :
RESULTS
Ginsenosides reduce the number of Fos-IR neurons increased by NS to tooth in the trigeminal spinal subnuclei and thalamus
To elucidate the nociceptive pathway of toothache, Fos-immunoreactive (IR) neurons were observed in the brain 1.5 h after noxious intrapulpal stimulation (NS) to the upper and lower incisor. Fos-IR neurons were absent or rare in brain regions of the rats without NS. After NS to tooth, Fos-IR neurons were bilaterally increased in dorsal superficial laminae of the trigeminal spinal subnucleus caudalis (Vc) and transitional region between Vc and subnucleus interpolaris (Vi) near the obex (Fig. 2). The most numerous labellings were found in the dorsomedial part, especially at 1 mm caudal to the obex in the Vc. However, trigeminal subnucleus oralis did not show Fos-IR neurons and trigeminal subnucleus interpolaris showed a few Fos-positive neurons (data not shown).
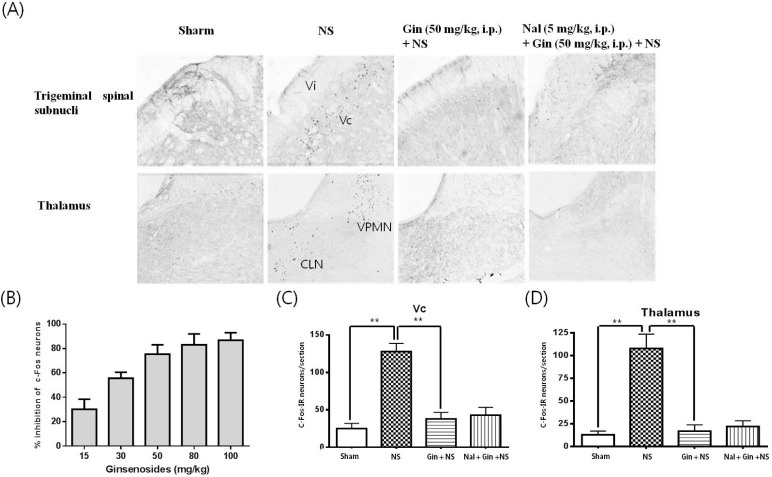 | Fig. 2Effects of ginsenosides on the number Fos-IR neurons increased by NS to tooth in the trigeminal spinal subnuclei and thalamus. (A) c-Fos immunoreactivity in trigeminal spinal subnuclei and thalamus. (B~D) Fos-IR neurons were increased in the trigeminal spinal subnucleus candalis (Vc) and transitional region between subnucleus interpolaris (Vi), thalamic ventral posteromedial nucleus (VPM) and centrolateral nucleus (CLN). 1.5 h after NS to the upper and lower incisor pulp with 2 M KCl. Ginsenosides reduced the number of Fos-IR neurons increased by NS to tooth in a dose-dependent manner and IC75 for ginsenosides is 50 mg/kg. Naloxon (Nal; 5 mg/kg, i.p.), an opioid antagonist, did not block the inhibitory effects of ginsenosides on the number of Fos-IR neurons increased by NS to tooth. Data represent as mean±SEM (n=5), **p<0.01.
|
Fos-IR neurons were enhanced by NS to tooth in the thalamic ventral posteromedial nucleus (VPMN), a specific sensory relay nucleus and centrolateral nucleus (CLN), a nonspecific sensory relay nucleus (Fig. 2). Intraperitoneal adiminstration of ginsenosides suppressed the number of Fos-IR neurons increased by NS to tooth in the trigeminal spinal subnuclei and thalamus in a dose-dependent manner and IC75 for ginsenosides is 50 mg/kg (Fig. 2). Naloxon, an opioid antagonist, was intraperitoneally injected 20 min prior to the noxious intrapulpal stimulation to examine whether the antinociceptive effects of ginsenosides are mediated through opioid system or not. Naloxone (5 mg/kg) had no blocking effects on the ginsenoside-induced suppressive effect for the number of Fos-IR neurons increased by NS to tooth (Fig. 2).
Ginsenosides rescue the cardiovascular responses elevated by NS to tooth
Mean BP and HR were recorded before and after intrapulpal noxious stimulation to the upper and lower incisor tooth under preadministration of ginsenoside or not. BP and HR were significantly increased immediately after noxious stimulation to tooth and returned close to the basal level since 20 min after noxious intrapulpal stimulation. Pretreatment of ginsenoside (50 mg/kg, i.p.) reduced arterial BP and HR increased by NS to the tooth (Fig. 3).
 | Fig. 3Effects of ginsenosides on the cardiovascular responses elevated by NS to tooth. The blood pressure (BP) and heart rate (HR) were increased immediately after NS to tooth. Ginsenosides (Gin; 50 mg/kg, i.p.) ameliorated the BP and HR increased by NS to tooth. Data represent as mean±SEM (n=12), *p<0.05.
|
Ginsenosides decrease the number of Fos-IR neurons increased by NS to tooth in central cardiovascular regulating areas
The NS to tooth augmented the number of Fos-IR neurons in the nucleus tractus solitaries (NTS) and rostral ventrolateral medullac (RVLM), central pressor areas (Fig. 4). Besides, the NS to tooth increased the number of Fos-IR neurons in the hypothalamic paraventricular nucleus (PVN) and supraoptic nucleus (SON), vasopressin and antidiuretic hormone-releasing regions (Fig. 5). Ginsenosides (50 mg/kg, i.p.) reduced the number of Fos-IR neurons increased by NS to tooth in NTS, RVLM, PVN and SON, but had no changes in BP and HR by its self (Fig. 4 and 5).
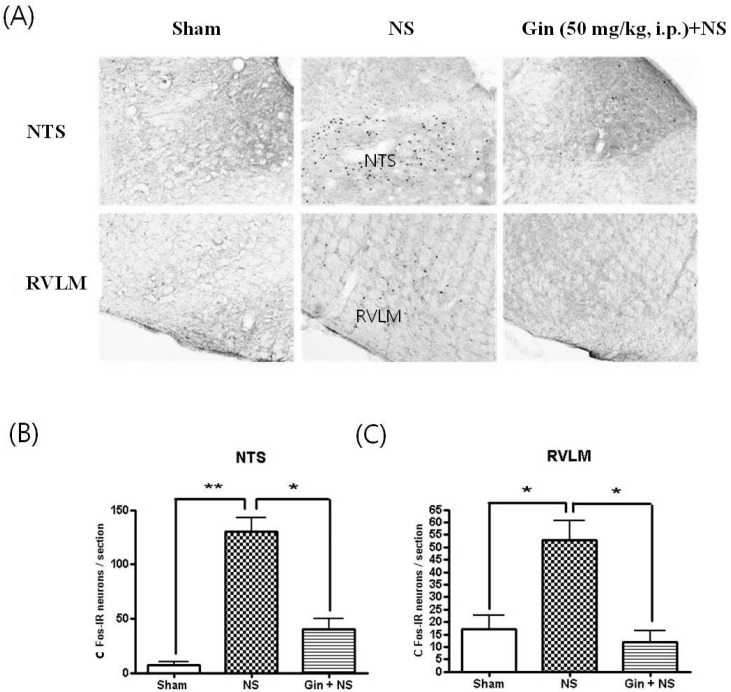 | Fig. 4Effects of ginsenosides on the number of Fos-IR neurons increased by NS to tooth in the nucleus tractus solitaries (NTS) and rostral ventrolateral medulla (RVLM). (A) c-Fos immunoreactivity in NTS and RVLM. (B, C) The NS to tooth caused a robust increment of Fos-IR neurons in the NTS and RVLM, central pressor regulating areas and ginsenosides (Gin; 50 mg/kg, i.p.) reduced the number of Fos-IR neurons increased by NS to tooth in the NTS and RVLM. Data represent as mean±SEM (n=5), *p<0.05, **p<0.01.
|
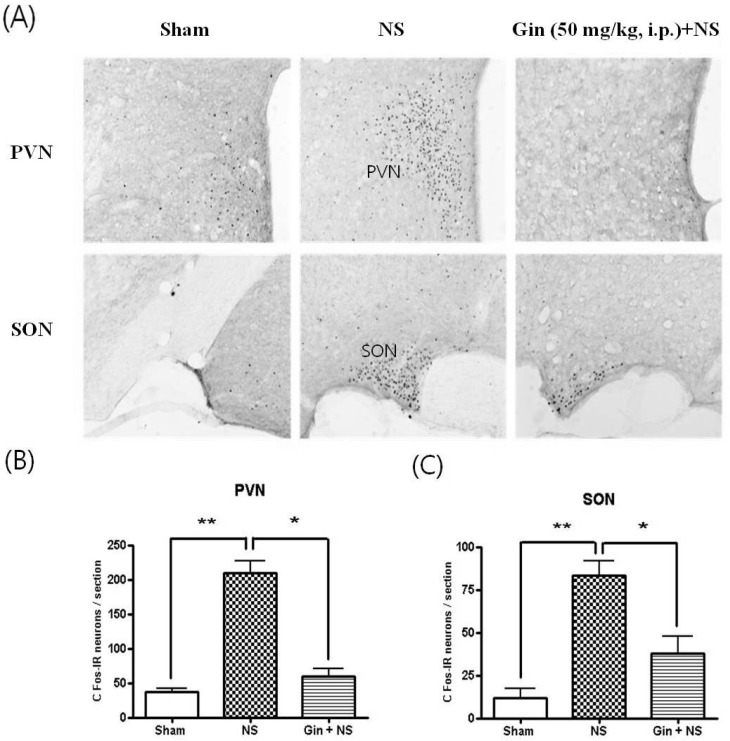 | Fig. 5Effects of ginsenosides on the number of Fos-IR neurons enhanced by NS to tooth in the hypothalamic paraventricular nucleus (PVN) and supraoptic nucleus (SON). (A) c-Fos immunoreactivity in PVN and SON. (B, C) Fos-IR neurons were significantly increased in the hypothalamic PVN and SON 1.5 h after NS to tooth and ginsenoside (Gin; 50 mg/kg, i.p.) reduced the number of Fos-IR neurons increased by NS to tooth. Data represent as mean±SEM (n=5), *p<0.05, **p<0.01.
|
Go to :
DISCUSSION
c-Fos mRNA expresses in post-synaptic neurones of central nervous system (CNS) a few minutes after peripheral noxious stimulation. c-Fos forms a part of the AP1 transcription factor complex which subsequently controls the expression of other genes associated with the short and long-term changes in neural excitability [12,13]. Subsequently, c-Fos protein can be found in CNS within 1~2 h of transcription. Even in the present experiment, c-Fos proteins were expressed in various CNS regions related to the tooth pain pathways 1.5 h after intrapulpal noxious stimulation.
In this study, c-Fos expression following the noxious intrapulpal stimuli was mainly observed in the superficial laminae (I, II) of Vc and the transition zone near to the obex between subnucleus caudalis and Vi. However, subnucleus oralis did not show c-Fos expression and a few c-Fos-positive neurons were observed only in subnucleus interpolaris. A pattern of Fos expression in this experiment was consistent with those evoked by pulpal heat stimulation or orofacial inflammation [1,14,15]. From the present results and some previous reports, it is speculated that the Vc and the Vi/Vc transitional regions play a pivotal role in tooth pulpal nociception. Besides, c-Fos protein was found in the thalamic VPM and CLN, a specific relay nucleus and nonspecific relay nucleus known as a receiving region of the second order neurons from spinal sensory nucleus. Taken together, it is concluded that nociceptive signals from tooth are conducted through trigemical Vc into the thalamus.
As a traditional herbal medicine, ginseng has been utilized to relieve a variety of pains such as chest pain, visceral pain and neuralgia. Ginsenoside, an active component of ginseng, has been known to relieve pain produced by noxious multiple stimuli previous in vivo animal model using behavioral test. However, the effects of ginsenoside on tooth pulpal pain model and its antinococeptive properties have not been studied. In the present study, intraperitoneal administration of ginsenoside reduced the number of Fos-IR neurons increased by NS to tooth in the trigeminal spinal subnuclei and thalamic nuclei expressed, and its suppressive effects on c-Fos were not blocked by naloxone, an opioid antagonist. These findings suggest that ginsenosides can produce analgesic effect on tooth pulpal pain and its antinociceptive mechanism seems unlikely to be mediated by opioid receptors. However, it is unclear how ginsenosides act to produce the antinociceptive effects through central or perpheral nervous system. One possible mechanism is that ginsenosides Rf inhibit voltage-dependent Ca2+ channels in sensory neurons and nociceptive neurons [9,16]. Another possibility is that ginsenoside may work by attenuating the responsiveness to glutamate/substance P of post-synaptic neurons in brain stem since intrathecal administration of ginsenosides attenuates the pain-related behavior produced by intrathecal injection of substance P of MNDA [17,18]. Therefore, it is speculated that ginsenoside may act on both pre- and post-synaptic sites at brain stem level.
On the other hand, orofacial pain has been well known to elicit the cardiovascular reflex, resulted in the arterial pressure and heart rate changes [2,3]. Even in the present study, the NS to tooth elevated arterial blood pressure and heart rate. However, central nervous mechanisms underlying the cardiovascular reflex evoked by intrapulpal pain have been unknown. Previous studies in the rabbit and cat have documented that the cardiovascular changes induced by electrical stimulation of tooth pulp are mediated in the brain stem because midbrain transection and spinal trigeminal tractomy of caudal medulla left the reflex responses intact [2]. In this study, the NS to tooth significantly increased the number of c-Fos expression in SON and PVN which release vasopressin and antidiuretic hormone (ADH), resulted in the elevation of blood pressure. Besides, noxious tooth stimulation also evoked c-Fos expression in nucleus tractus solitarius (NTS) and rostral ventrolateral medulla (RVLM), central pressor centers. It was recently shown that neurons in the hypothalamic SON and PVN project to RVLM which is known as final common pathway in regulating the sympathetic nerve activity to accomplish the baroreflex [19,20]. In addition, it was reported that trigeminal sensory nucleus have efferent projections to the hypothalamic nucleus and NTS. Thus, it is likely that there is a functional linkage between the trigeminal sensory nucleus and thalamic nucleus or NTS as well as between the hypothalamus and RVLM. From these results, it is speculated that tooth pulpal pain evoked the cardiovascular reflex mediated by SON, PVN, NTS and RVLM of which mechanisms will be further needed to study.
Taken together, the present study showed that intraperitoneal administration of ginsenosides ameliorated the arterial blood pressure and heart rate elevated by NS to tooth. Moreover, ginsenosides (50 mg/kg, i.p.) decreased the number of c-Fos-IR neurons increased by NS in NTS, RVLM, PVN and SON, the central cardiovascular regulating centers. These results demonstrated that ginsenosides can rescue the cardiovascular responses altered by NS to tooth. However, it is not still clear that ginsenosides directly act in the cardiovascular system or indirectly act through the blocking of pain pathway since some previous reports have showed that ginsenosides decrease blood pressure via its direct action in cardiovascular system [21,22].
Go to :
 XML Download
XML Download