

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Extracellular pH and systemic acid-base status are critical for normal organ and cellular function. The kidney controls and maintains systemic acid-base status by intricately linked mechanisms: reabsorption of filtered HCO3-, excretion of acid, and de novo generation of ammonium and bicarbonate [1]. In these processes, acid-base transporters are involved. A variety of mediators of transport regulation in the kidney work by both short term and long term actions. Long term regulations are associated with increases in abundances of transporter proteins, while short term regulations are associated with increased trafficking as well as post-translational modification of the transport proteins [2].
Approximately 70% of filtered sodium is reabsorbed in the proximal tubule. A large portion of the apical component of sodium, proton and ammonium transport is mediated by the type 3 Na+/H+ exchanger (NHE3) [3]. In the basolateral membrane, type 1 Na+/HCO3- cotransporter (NBC1) carries sodium against an electrochemical gradient owing to its coupling with bicarbonate anion [4]. The ultimate fine tuning of renal acid/base transport occurs in the cortical collecting duct. Intercalated cells, which make up a third of cells of the cortical collecting duct, are responsible for acid-base transport [1]. Type A intercalated cells secrete H+ into the urine via a vacuolar H+-ATPase which is located to the apical plasma membrane and apical vesicles, and reabsorb HCO3- to the blood through anion exchanger-1 (AE1) localized to the basolateral plasma membrane [5,6]. Type B intercalated cells secrete HCO3- in exchange for Cl- across the apical membrane through the novel anion exchanger, pendrin [7].
In response to an acid load, NH4+ excretion begins to increase within 2 h in human study, due mostly to the formation of more acid urine which increases the efficiency of NH3 secretion into the medullary collecting tubule [8]. Total NH4+ excretion reaches its maximum level at 5 to 6 days, a time at which there is an elevation in both glutamine uptake by the kidney and tubular NH4+ production [8]. Animal models provide confirmation of this sequence. Metabolic acidosis was more pronounced in animals receiving NH4Cl for 2 days, as evident from blood pH and plasma bicarbonate concentrations, indicating partial adaptation of these animals to acid base disturbances after 7 days of acid loading [9]. These findings may suggest that there are dynamic changes in acid secretion at different stages (i.e., developmental stage and an adaptive or compensatory stage) of acidosis. However, the molecular basis for the dynamic changes of acid-base transporters in the kidney remains undefined. The present study was aimed at examining the changes in expression of key renal acid-base transporters in response to short term and long term acid loading in rats.
Go to : 
METHODS
Animals
Male Sprague-Dawley rats weighing 180~200 g were used. Rats were kept in a temperature-controlled room regulated on a 12:12-h light-dark cycle. The study was approved by the Ethics Committee of Chonnam National University Medical School.
Metabolic studies
Protocol 1: acute NH4Cl loading experiment
Metabolic acidosis was induced by intra-gastric gavage feeding of NH4Cl (21 mmol/kg/day) for 2 days [10]. The rats were given morning and evening doses (10.5 mmol/kg) for 2 days and a final dose on the morning of day 3 just prior to anesthesia. Rats were maintained in the metabolic cages to allow urine collections for the measurements of pH, Na+, Cl- and creatinine. Rats were anesthetized by intraperitoneal injections of ketamine and xylazine, and heparinized arterial blood was collected from the arterial puncture, and analyzed immediately for pH, pCO2, HCO3-, Na+ and creatinine. The kidneys were removed and processed for semiquantitative immunoblotting.
Protocol 2: chronic NH4Cl loading experiment
Another set of NH4Cl loading rats and control rats was made. Control and NH4Cl loading rats were chosen randomly, and all were provided with a gelled agar (1%) diet. By using this gelled diet, a daily, fixed amount of water [37 ml/220 g body wt/day] and regular rat chow (15 g/220 g BW/day) were given to each rat from the time it was placed in metabolic cages throughout the study period. The rats were fed once daily and ate all of the offered food during the course of the day. To investigate the effects of chronic NH4Cl loading, 34 mmol/kg/day of NH4Cl were included in the gelled diet and given for 7 days [11]. Control rats received the same gelled diet but without the NH4Cl.
Semiquantitative immunoblotting
The dissected cortex/outer stripe of outer medulla (OSOM) and inner stripe of outer medulla (ISOM) were homogenized in ice-cold isolation solution containing 0.3 M sucrose, 25 mM imidazole, 1 mM ethylenediaminetetraacetic acid (EDTA), 8.5 µM leupeptin, 1 mM phenylmethylsulfonyl fluoride (PMSF), with pH 7.2. The homogenates were centrifuged at 1,000 g for 15 min at 4℃ to remove whole cells, nuclei, and mitochondria. The total protein concentration was measured. All samples were adjusted with isolation solution to reach the same final protein concentrations and solubilized at 65℃ for 15 min in sodium dodecyl sulfate (SDS)-containing sample buffer and then stored at -20℃. To confirm equal loading of protein, we stained an initial gel with Coomassie blue. SDS-polyacrylamide gel electrophoresis was performed on 9 or 12% polyacrylamide gels. Subsequently, immunoblotting was performed as described previously [12]. The proteins were transferred by gel electrophoresis (Bio-Rad, Mini Protean II; Hercules, CA, USA) onto nitrocellulose Membranes (Amersham Pharmacia Biotech, Hybond ECL RPN3032D; Little Chalfont, UK). The blots were subsequently blocked with 5% milk in PBS with tween 20 (80 mM Na2HPO4, 20 mM NaH2PO4, 100 mM NaCl, 0.1% Tween 20, pH 7.5) for 1 h and incubated overnight at 4℃ with primary antibodies, followed by incubation with secondary anti-rabbit (Cell signaling, Boston, USA) or anti-mouse (Cellsignaling) horseradish peroxidase-conjugated antibodies. The labeling was visualized by an enhanced chemiluminescence system.
The primary antibodies to Na+/H+ Exchanger 3 (NHE3), Na+-HCO3-cotransporters (NBC1), Anion Exchanger-1 (AE-1, Alpha Diagnostic, San Antonio, USA), Na+ -K+ ATPase (Cell signaling, Boston, USA) and H+-ATPase (Santa Cruz, Bergheimer, Germany) were commercially obtained.
Statistical analyses
Results are expressed as mean±SEM. The statistical significance of differences between the groups was determined using an unpaired t-test. Differences with values of p<0.05 were considered significant.
Go to :
RESULTS
Blood and urine parameters
Table 1 shows blood and urine parameters from control and 2 days of NH4Cl-loaded rats. Arterial blood gas analysis showed decreased bicarbonate levels (19.86±0.49 vs. 28.60±1.92 mEq/l, p<0.05) in NH4Cl-loaded rats compared with controls. Accordingly, blood pH (7.35±0.04 vs. 7.47±0.02, p<0.05) and pCO2 (37.6±0.55 vs. 41.2±2.74 mmHg, p<0.05) levels were decreased. Serum sodium level (135.4±1.81 vs. 139.2±2.58 mEq/l, p<0.05) was decreased in NH4Cl-loaded rats. On the other hand, creatinine level was not different between two groups. Urine pH (6.4±0.1 vs. 7.0±0.39, p<0.05) was decreased, while urine chloride (11.2±2.30 vs. 3.1±0.62 mEq/day, p<0.05) was increased in NH4Cl-loaded rats.
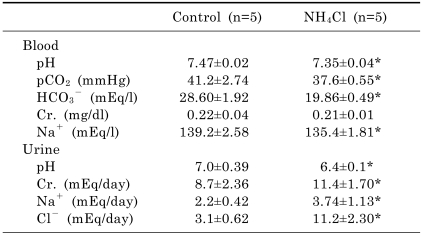
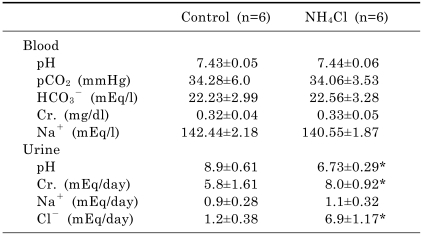
Table 2 shows blood and urine parameters from control and 7 days of NH4Cl-loaded rats. Arterial blood gas analysis revealed that blood pH, pCO2 and HCO3- levels were not different between two groups. On the other hand, urine pH (6.73±0.29 vs. 8.9±0.61, p<0.05) was markedly decreased, while urine chloride level was increased (6.9±1.17 vs. 1.2±0.38 mEq/day, p<0.05) in NH4Cl-loaded rats.
Alteration of acid-base transporters
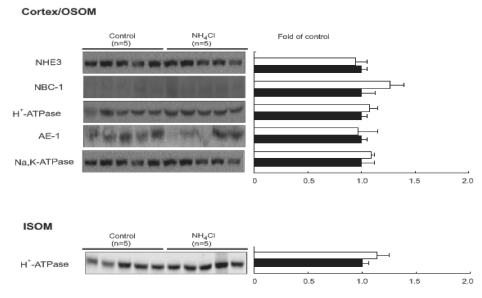
We investigated the expression of key renal acid-base transporters from control and 2 days of NH4Cl-loaded rats. The protein expression of NHE3, NBC1, H+-ATPase, AE1 and Na+ -K+ ATPase, was not different in the kidney cortex/OSOM from NH4Cl-loaded rats compared with controls. The protein expression of H+-ATPase was not changed in ISOM (Fig. 1).
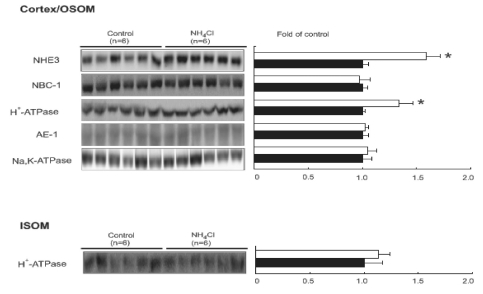
Subsequently, the key renal acid-base transporters from control and 7 days of NH4Cl-loaded rats were investigated. The protein expression of NHE3 and H+-ATPase was increased in cortex/OSOM from NH4Cl-loaded rats compared with that in controls, while NBC1, AE1 and Na+ -K+ ATPase, expressions were not different. The protein expression of H+-ATPase was not changed in ISOM (Fig. 2).
Go to :
DISCUSSION
In the present study, we investigated the time course changes of urinary acidification in response to ammonium chloride loading, and showed the changes of key acid-base transporters in the kidney. To induce metabolic acidosis and compare adaptive changes in metabolic and transport pathways, NH4Cl was loaded for 2 days and 7 days with reference to other experiments in rats [13]. Unlike the NH4Cl loaded mice, NH4Cl loaded rats recovered from acidosis on day 7, which may reflect species differences [14,15].
The provision of 2 days of NH4Cl caused a significant metabolic acidosis manifested by decreased serum bicarbonate and blood pH. Accordingly, urine pH was decreased, while urine chloride excretion was substantially higher in NH4Cl loaded rats than in controls suggesting the higher chloride intake in NH4Cl loaded rats. In contrast, blood pH and bicarbonate levels in chronic ammonium chloride loaded rats were comparable with controls. These findings may suggest that there are subsequent changes in response to acid loading at different stages. It should be emphasized that the differences found in acute and chronic acid loading support the view that dynamic changes occur that likely represent changes in the onset of acidosis or represent compensatory changes.
The apical membrane Na+/H+ antiporter plays a important role in proximal tubular transepithelial NaHCO3 and NaCl transport, and in thick ascending limb NaHCO3 absorption [16]. Several experimental evidences indicated that the Na+/H+ antiporter isoform, NHE3, encodes a major component of apical membrane Na+/H+ antiporter activity in the proximal tubule and thick ascending limb [17,18]. Our data demonstrated that NHE3 abundance was not changed in response to acute acid loading, which coincided with the development of metabolic acidosis. On the other hand, protein expression of NHE3 was increased in chronic acid loading associated with recovery from acidosis, which suggest adaptive renal response to sustained increment in acid load. These finding were consistent with previous studies, which showed effect of acidosis on NHE3 abundance was beginning at 3 days and became significant at 7 days [19]. Basolaterally expressed Na-HCO3- cotransporter, NBC1, in the proximal tubule also plays an important role in mediating electrogenic HCO3- efflux [20-22]. Several NBC isoforms have been identified and have been studied for potential role in response to chronic metabolic acidosis. Kwon et al. showed increased protein abundance NBC isoforms in response to chronic metabolic acidosis, which is confined to NBCn1 and NBC3 but not NBC1 [23]. Our result is also consistent with changes of NBC isoforms in response to chronic acid loading, although analysis was limited to NBC1.
The collecting duct is the final site for urine acid-base regulation. The collecting duct is composed of a "tight" epithelium that can generate steep pH gradients and thus plays a major role in the excretion of net acid. Intercalated cells in the collecting duct secrete H+ into the urine via a vacuolar H+-ATPase which is located to the apical plasma membrane and apical vesicles. In the present study, we showed that H+-ATPase abundance was not changed in response to acute acid loading, and increased in chronic acid loading. These findings suggest that adapted activation of H+-ATPase in response to metabolic acidosis might counteract the development of metabolic acidosis. In addition to apical membrane H+-ATPase, reabsorb HCO3- to the blood through basolateral membrane localized AE1 plays a role in acid-base homeostasis. Several lines of evidence indicated that AE1 also provide adaptive mechanism in response to chronic metabolic acidosis [24,25]. In contrast, our data showed no difference in the abundance of AE1 in both acute and chronic acid loading. In the present study, on day 7, serum HCO3- level returned to normal reference range, which may suggest the possible influence of pH changes on AE1 abundance. Purkerson JM et al. showed that adaptive changes of AE1 rapidly reversed after recovery from acidosis [25].
The present study has several limitations. First, our analysis confined to altered expression of various transporter. In addition to these changes, regulation of phosphorylation and proliferation of acid-secretory cells also have been proposed for adaptive mechanism in response to chronic acid loading [26,27]. These physiologic responses also provided a comprehensive view for understanding renal adaptive response to metabolic acidosis, which was not evaluated in our experiment. Second, we evaluated altered protein expression in response to acid loading, which was measured by semiquantitative immunoblotting. In addition to altered protein expression, altered regulation of gene expression was observed during metabolic acidosis, which results from activation of transcription and/or mRNA stabilization [2,28]. Moreover, targeting of various transport in the tubular segment also play an important role in response to acid loading, which can be confirmed by immunohistochemical staining. Unfortunately, our study does not evaluate all these factors, and provided limited insight for the understanding of renal adaptive response in chronic metabolic acidosis. Despite of these limitations, our data demonstrated the time course change of systemic and urinary acidification in response to ammonium chloride loading, and elucidated the role of various transporters in renal tubular epithelium by observing the changes of protein expression.
In conclusion, our experiments suggest that not fully adapted activation of acid-base transporters in the kidney may contribute to the development of metabolic acidosis in response to acute NH4Cl loading. The subsequent increased expression of NHE3, H+-ATPase in the kidney may play a role in promoting acid secretion in the later stage of acid loading, which counteract the development of metabolic acidosis.
Go to :
 XML Download
XML Download