

 PDF
PDF ePub
ePub Citation
Citation Print
Print


ABBREVIATIONS
PA
Phellodendron amurense
BER
berberine
AD
Alzheimer's disease
ACh
acetylcholine
SCO
scopolamine
CNS
central nervous system
IL-1β
interleukin-1β
TNF-α
tumor necrosis factor-α
PAT
passive avoidance test
MWM
Morris water maze
TA
tacrine
ChAT
Cholineacetyltransferase
AchE
Acethylcholinesterase
PBS
phosphate-buffered saline
ABC
avidin-biotin-peroxidase complex
DAB
3,3'-diaminobenzidine
ELISA
Enzyme-Linked Immunosorbent Assay
BDNF
Brain-derived neurotrophic factor
CREB
cAMP-response element-binding protein
COX-2
cyclooxygenase-2
RT-PCR
reverse transcription-polymerase chain reaction
GAPDH
glyceraldehyde-3-phosphate dehydrogenase
ANOVA
analysis of variance
NF-κB
nuclear factor-kappaB
PGE2
prostaglandins E2
INTRODUCTION
Alzheimer's disease (AD), which is characterized by a progressive decline in cognitive functioning due to degeneration of the cholinergic nervous system and neuronal dysfunction, is one of the most common forms of dementia in the aging population [1]. In discussions of the neuropathological features of memory loss and cognitive dysfunction, as observed primarily, in patients with AD, cholinergic deficits and neuronal dysfunction have been considered a primary causes [2]. Accordingly, various cholinergic drugs have been approved to treat AD, and they exert their therapeutic effects by counteracting acetylcholine (ACh) deficits and consequently enhancing the ACh levels in the brain [3].
In the present study, intraperitoneal (i.p.) administration of the muscarinic antagonist scopolamine (SCO), a blocker of muscarinic ACh receptors, was exploited as a pharmacological model for AD in rats. SCO induces dysregulation of the cholinergic neuronal pathway and memory circuits in the central nervous system (CNS), resulting in serious impairments in learning, acquisition, and short-term retention of spatial memory tasks [4]. Because such lesions in cholinergic neuronal circuits result in decreased ACh release and subsequent dysfunction in learning and memory, the SCO-induced amnesic model has been widely used to provide a model system of memory dysfunction to screen for pharmacological agents with memory-enhancing activities [5].
Inflammation, as well as cholinergic neuronal degeneration, may play an important role in the pathogenesis of the degenerative changes and cognitive impairment associated with AD. Proinflammatory cytokines, such as interleukin-1β (IL-1β) and tumor necrosis factor-α (TNF-α) are upregulated in the AD brain [6]. These inflammatory cytokines may play a role as neurotoxic agents in the pathological cascade of AD [7].
Many studies have reported that the use of herbal medicine or natural products in treating Alzheimer type-dementia patients improved their memory related symptoms [8,9]. However, there has been little scientific evidence reported regarding their effectiveness or mechanisms of their action.
Phellodendron amurense (PA) has been widely used in Korean traditional medicine to treat bacterial infection of the respiratory system and chronic inflammatory diseases [10]. PA and its major alkaloid compound, berberine (BER), have multiple pharmacological actions and produce a variety of biological effects in the CNS [11]. In particular, BER has been reported to improve diabetes-induced memory impairment and to reduce oxidative stress [12]. Several studies have shown that BER was effective in ameliorating spatial memory impairment and decreasing the expression of IL-1β in a rat model of AD [13], suggesting that the anti-inflammatory effects of PA and BER may result from their ability to inhibit the degeneration of cholinergic neurons, thereby alleviating deficits in spatial learning ability in the memory-impairment animal model [14].
The aim of the present study was to evaluate the ability of PA and BER to improve learning and memory in rats exposed to repeated SCO-induced memory deficits as measured by performance on the passive avoidance test (PAT) and the Morris water maze (MWM) test. Moreover, we examined how these effects were related to the cholinergic system and to anti-inflammatory effects to determine the neural mechanisms underlying the memory-enhancing activity of PA and its main alkaloid compound, BER.
Go to : 
METHODS
Animals
Adult male Sprague-Dawley (SD) rats weighing 260~280 g were obtained from Samtako Animal Co. (Seoul, Korea). The rats were housed in a limited-access rodent facility with up to five rats per polycarbonate cage. The room controls were set to maintain the temperature at 22±2℃ and the relative humidity at 55±15%. Cages were lit by artificial light for 12 h each day. Sterilized drinking water and standard chow diet were supplied ad libitum to each cage during the experiments. The animal experiments were conducted in accordance with the Guide for the Care and Use of Laboratory Animals (NIH Publications No. 80-23), revised in 1996, and were approved by the Kyung Hee University Institutional Animal Care and Use Committee. All animal experiments began at least 7 days after the animals arrived.
Main reagent
Scopolamine hydrobromide, tacrine (TA) and BER were obtained from standard commercial suppliers (Sigma-Aldrich Chemical Co., St Louis, MO, USA). PA, used in this study, was purchase from an oriental drug store (Health maximum Co., Jecheon, Korea). A voucher specimen of PA has been deposited at the herbarium located at the College of Oriental Medicine, Kyung Hee University (Code number KH-PA01 for PA), according to described previous study [15]. One hundred grams of PA were cut into small pieces and extracted three times with 4 l of 85% methanol by sonication in a reflux condenser for 24 h at room temperature (25±2℃). The extracted solutions were combined, filtered through Whatman No. 1 filter paper, concentrated using a rotary vacuum evaporator (EYELA CCA-1110, Tokyo Rikakikai Co., Tokyo, Japan) to obtain concentrated extracts, and then lyophilized (EYELA FD-800). The final yield of PA in a powder form was 13.7% (w/w).
Experimental groups
To develop learning and memory deficits, male rats were intraperitoneally injected at 2 mg/kg body weight with SCO, dissolved in physiological saline solution, once a day for 14 days. Normal animals received saline instead of SCO as a vehicle. Different rats in an experimental group were subjected to either behavioral testing or immunohistochemistry. The rats were randomly divided into seven groups of seven individuals as follows: normal group (SAL group, n=7), the saline-induced plus 200 mg/kg PA-treated group (PA group, n=7), the SCO-induced and saline-treated group (SCO group as a control, n=7), the SCO-induced plus 20 mg/kg BER-treated group (SCO+BER20 group, n=7), the SCO-induced plus 100 mg/kg PA-treated group (SCO+PA100 group, n=7), the SCO-induced plus 200 mg/kg PA-treated group (SCO+PA200 group, n=7), and the SCO-induced plus 0.2 mg/kg TA-treated group (SCO+TA group, n=7). TA, a centrally acting cholinesterase inhibitor, was used as a positive control. The rats were intraperitoneally administrated with PA and BER once a day for 14 days, and PA and BER were dissolved in 0.9% physiological saline. Thirty minutes after PA and BER administration, all rats except SAL group were received the SCO injection. When starting 9th day after SCO injection, rats performed to take the MWM task. The experimental schedule of all drug administration and behavioral tests are shown in Fig. 1.
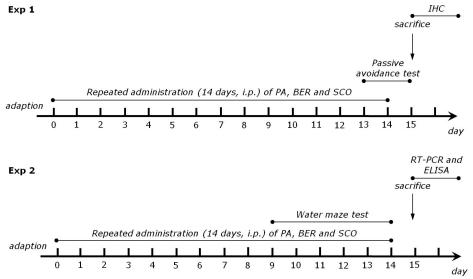 | Fig. 1Experimental schedules of scopolamine-induced spatial memory impairments in the rats. Exp 1 was designed to explore the efficacy of PA and BER administration for amelioration of memory impairment in an animal model using a passive avoidance test (PAT) and immunohistochemistry. Exp 2 was designed to explore the efficacy of PA and BER administration for amelioration of memory impairment in an animal model using the Morris water maze (MWM) test, ELISA and RT-RCT analyses.
|
Passive avoidance test
All animals were subjected to a passive avoidance test. The test was basically performed according to the step-through method described previously [16]. The Gemini Avoidance System (SD Instruments., San Diego, CA, USA) was used for this experiment. Basically, the step-through passive avoidance apparatus consists of a two-compartment acrylic box with a lightened compartment connected to a darkened one by an automatic guillotine door. Electric shock was delivered to the grid floor of both compartment, made of stainless steel rods (3 mm diameter) spaced 1 cm apart, by an isolated shock generator (Behbood Pardaz Co., Ghaem, Iran). First, rats were taken trials to acquisition test in the apparatus. In this trial, rats were placed in a lightened compartment for 300 s, and then the guillotine door was opened. Rats have native preference to the dark environment. Immediately upon entering the dark compartment, the door was closed. Acquisition test was recorded the latencies times for entering the dark compartment. After 30 min, rats were again placed in the lightened compartment. Rats had spontaneously entered the dark compartment, the guillotine door was closed and a mild electrical shock (0.5 mA) was applied for 3 s. Exactly 24 h after the acquisition trial for training, the retention test was performed. The rat was again placed in the lightened compartment and the guillotine door was opened. The retention test was measured the latencies times for entering the dark compartment in a same method with acquisition test. The maximum entry latency allowed in the retention test was 180 s [17].
Morris water maze test
1. Morris water maze apparatus
The MWM test was performed using a polypropylene circular pool (painted white internally, 2.0 m in diameter and 0.35 m high). The pool contained water maintained at a temperature of 22±2℃. The water was made opaque by adding 1 kg of skim milk powder. During the MWM test, a platform 15 cm in diameter was located 1.5 cm below the water in one of four sections of the pool, approximately 50 cm from the sidewalls. The pool was surrounded by many cues external to the maze. The pool was divided into four quadrants of equal area. A digital camera was mounted to the ceiling above the pool and was connected to a computerized recording system equipped with a tracking program (S-MART: PanLab Co., Barcelona, Spain), which permitted on- and off-line automated tracking of the paths taken by the rats.
2. Hidden platform trial for acquisition test
The MWM test was initiated on the 9th day after the PA, BER and SCO administration commenced. The animals received three trials per day. The rats were trained to find the hidden platform, which remained in a fixed location throughout the test. The trials lasted for a maximum of 180 s, and the time it took to find the submerged platform was recorded each time. The animals were tested three trials per day for 5 days, and they received a 60-s probe trial on the sixth day. Finding the platform was defined as staying on it for at least 4 s before the acquisition time of 180 s ended. If the rat failed to find the platform in the allotted time in first trial of hidden platform test, those rats should placed onto the platform for 20 s and assigned a latency of 180 s. Between one trail and the next, water was stirred to erase olfactory traces of previous swim patterns. The entire procedure took sixth consecutive days, and each animal had three training trials per day, with a 30- to 40-min intertrial interval.
3. Probe trial for retention test
For the probe trial, each rat was placed into the water diagonally from the target quadrant, and for 60 s, was allowed to search the water from which the platform had been removed. The probe trial was expressed by the ratio of the time spent for searching the platform in the target quadrant to total duration spent for swimming in the pool.
Cholineacetyltransferase (ChAT) and Acethylcholinesterase (AchE) immunohistochemistry
For immunohistochemical studies, the animals were deeply anesthetized with sodium pentobarbital (80 mg/kg, by intraperitoneal injection) and perfused through the ascending aorta with normal saline (0.9%) followed by 300 ml (per rat) of 4% paraformaldehyde in 0.1 M phosphate-buffered saline (PBS). The brains were removed, post-fixed over-night, and cryoprotected with 20% sucrose in 0.1 M PBS at 4℃. Coronal sections 30 µm thick were cut through the septal region and hippocampus using a cryostat (Leica CM1850; Leica Microsystems Ltd., Nussloch, Germany). The sections were immunostained for ChAT expression using the avidin-biotin-peroxidase complex (ABC) method. Briefly, the sections were rinsed three times for 5 min each in PBS and then incubated with primary rabbit anti-ChAT antibody (1:2,000 dilution; Cambridge Research Biochemicals Co., Bellingham, UK), or goat anti-AchE antibody (1:2,000 dilution; Santa Cruz Biotechnology Inc., CA, USA) in PBST (PBS plus 0.3% Triton X-100) for 72 h at 4℃. Both secondary antibodies were obtained from Vector Laboratories Co. (Burlingame, CA, USA) and diluted 1:200 in PBST containing 2% normal serum. To visualize immunoreactivity, the sections were incubated for 90 min in ABC reagent (Vectastain Elite ABC kit; Vector Labs. Co., Burlingame, CA, USA), and incubated in a solution containing 3,3'-diaminobenzidine (DAB; Sigma) and 0.01% H2O2 for 1 min. Images were captured using the AxioVision 3.0 imaging system (Carl Zeiss, Inc., Oberkochen, Germany) and processed using Adobe Photoshop (Adobe Systems, Inc., San Jose, CA, USA). The sections were viewed at 200×magnification, and the numbers of cells within 100×100-µm2 grids were counted by observers blinded to the experimental groups. Medial septum and hippocampal area cells were obtained according to the stereotactic atlas of Paxinos and Watson [18]. The cells were counted in three sections per rat within the hippocampal area.
Enzyme-Linked Immunosorbent Assay (ELISA) in plasma
After SCO injection for 14 days, IL-1β and IL-6 concentration in blood was determined. For this study, the unanesthetized rats were rapidly decapitated, and the blood was quickly collected via the abdominal aorta. Homogenization was carried out on ice using a tissue homogenizer and incubated for 1 min at 4℃ with shaking. Homogenates were centrifuged and supernatants were collected. Protein concentrations were estimated by the procedure of Taylor et al. [19] with BSA as the standard. The IL-1β and IL-6 concentration was measured by a competitive enzyme-linked immunoassay (ELISA) using a rabbit polyclonal IL-1β and IL-6 antibody (IL-1β and IL-6; R&D Systems, Minneapolis, MN, USA) according to the manufacturer's protocol. Samples (or standard) and conjugate were added to each well, and the plate was incubated for 1 h at room temperature without blocking. After wells were washed several times with buffers and proper color developed, the optical density was measured at 450 nm using an ELISA reader (MutiRead 400; Authos Co., Vienna, Austria).
Total RNA preparation and RT-PCR analysis
The hippocampus from each of four rats in each group was isolated. After decapitation, the brain was quickly removed and stored at -80℃ until use. The total RNA was isolated from the brain sample using a TRIzol® reagent (Invitrogen Co., Carlsbad, CA, USA) and RNA was extracted according to the supplier's instruction. Complementary DNA was synthesized from total RNA with a reverse transcriptase (Takara Co., Shiga, Japan). Brain-derived neurotrophic factor (BDNF), cAMP-response element-binding protein (CREB), IL-1β, TNF-α, and cyclooxygenase-2 (COX-2) mRNA expression levels were determined by the reverse transcription-polymerase chain reaction (RT-PCR). RT-PCR was performed using a PTC-100 programmable thermal controller (MJ Research, Inc., Watertown, MA, USA). All primers were designed using published mRNA sequences and primer design software (Primer 3; The Whitehead Institute for Biomedical Research, Cambridge, MA, USA; www.genome.wi.mit.edu) offered through their website. The PCR products were separated on 1.2% agarose gels and stained with ethidium bromide after which the density of each band was evaluated using an image-analyzing system (i-Max™, CoreBio System Co., Seoul, Korea). Complementary DNA expression levels were determined by calculating the relative density of each BDNF, CREB, IL-1β, TNF-α, and COX-2 band to glyceraldehyde-3-phosphate dehydrogenase (GAPDH).
Statistical analysis
All measurements were performed by an independent investigator blinded to the experimental conditions. Results in figures are expressed as mean±standard error of means (SE). Differences within or between normally distributed data were analyzed by analysis of variance (ANOVA) using SPSS (Version 13.0; SPSS, Inc., Chicago, IL, USA) followed by Tukey's post hoc test. Statistical significance was set at p<0.05.
Go to :
RESULTS
Effect of PA and its main alkaloid compound, BER on the passive avoidance test
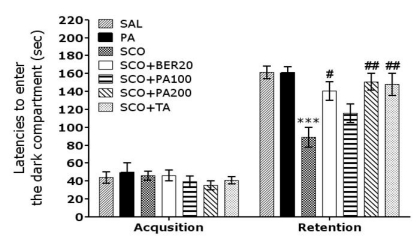
To determine whether PA or BER promotes recovery of memory dysfunction, PA or BER was administered to rats with SCO-induced impairment of memory, and their memory and cognitive functions were examined by the PAT (Fig. 2). The rats in all groups were confirmed to have no physiological defect (i.e., motor function defect) or intrinsic cognitive impairment prior to SCO-induced impairments established through acquisition trials without electric challenge in the PAT. No significant difference was found between the groups in the time required for acquisition trials, as indicated by the latencies for entering the dark compartment. After acquisition, the effect of PA or BER on retention latency, indicated by the latency times for entering the dark compartment, was measured 24 hours after applying electric shock in the dark room in the PAT. Regarding retention, rats in the SCO+BER20 group (p<0.05) and the SCO+PA200 group (p<0.01) were shown to have significantly increased latencies for entering the dark compartment compared with the SCO group. This study indicates that SCO injection severely impaired long-term memory, while the PA and BER treatment significantly attenuated the SCO-induced memory deficit in the PAT test. It also indicated that the recovery of memory impairment of rats in the 200 mg/kg PA-treated group was better than those rats in the 20 mg/kg BER-treated group, and almost compatible with the rats in the 0.2 mg/kg TA-treated group.
Effect of PA and its main alkaloid compound, BER on the water maze test
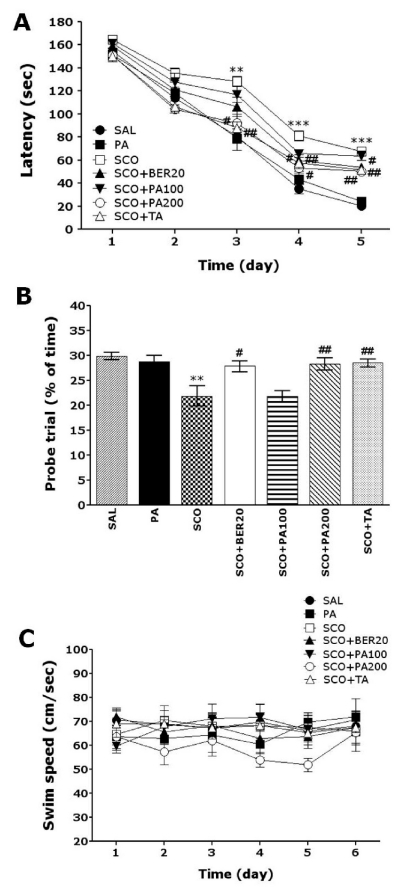
The effect of PA or BER treatment on the swimming time to reach the submerged platform is illustrated in Fig. 3. The SAL group rapidly learned the location of the submerged hidden platform and reached it within 20 s on day 5 of the trials. The SCO group showed a marked retardation in escape latency, probably due to memory deficits resulting from SCO-induced impairment of learning and memory. Analysis of escape latency revealed that rats in both the SCO+BER20 group and the SCO+PA200 group had significantly reduced swimming latency as compared with those in the SCO group (SCO+BER20 group: p<0.05 on day 4 and 5, and SCO+PA200 group: p<0.05 on day 3, p<0.01 on day 4 and 5; Fig. 3A). To examine the spatial memory of rats, the performance in the probe trial on day 6 was analyzed by comparing the percentage of time spent swimming to the expected position of platform (Fig. 3B). The times spent swimming around was significantly reduced in the rats that swam to the target area where the platform had been located. The repeated administration of SCO severely impaired spatial performance in the MWM test (p<0.01). Rats in the 20 mg/kg BER-treated (p<0.05) group and the 200 mg/kg PA-treated (p<0.01) group spent more time around the platform area than did those in the SCO group. The SCO group was not significantly different from other groups in terms of mean swimming speed, as calculated by dividing the total swim distance by the latency (Fig. 3C). Total distance traveled in each group was closely associated with escape latency in this task (data not shown). Based on these results, the 20 mg/kg of BER-treated and 200 mg/kg of PA-treated rats showed greater improvements in acquisition during the hidden platform trial and reached the platform quicker than the SCO-treated rats. PA and BER treatment significantly attenuated the SCO-induced deficit of learning and memory demonstrated in the water maze. Thus, PA-treated rats and BER-treated rats showed a significant amelioration in the memory retention test because they spent more time in the quadrant where the platform was formerly located and swam over the former location of the platform more frequently. It also indicated that the swimming latency of spatial memory impairment in rats receiving 200 mg/kg PA administration was better than those rats receiving 20 mg/kg BER administration, and almost compatible with the rats receiving 0.2 mg/kg TA administration.
Effect of PA and its main alkaloid compound, BER on septo-hippocampal choline acetyltransferase
Following the behavioral tasks, brain tissue samples from the subjects were analyzed using immunohistochemistry to investigate the effect of PA or BER administration on neurons loss due to SCO-induced memory impairment. ChAT immunoreactivity analysis in the septo-hippocampal cholinergic neurons areas are shown in Fig. 4. The brains of the SCO group showed significant neurons loss in the medical septum and hippocampal regions of septo-hippocampal cholinergic neurons, as compared to the SAL group (p<0.001; Fig. 5A). The number of ChAT-immunoreactive neurons significantly increased in the medical septum region in the SCO+BER20 group (p<0.01) and the SCO+PA200 group (p<0.01), as compared to the SCO group. Also, the number of ChAT-immunoreactive neurons significantly increased in hippocampal region in the SCO+BER20 group (p<0.05) and the SCO+PA200 group (p<0.05), as compared to the SCO group. Losses of ChAT-immunoreactivity in SCO-induced memory impairment rats were significantly restored by the PA and BER administration, and the number of ChAT-immunopositive neurons was closely similar to that of the control group. It also indicated that the ChAT activity in the septo-hippocampal cholinergic neurons in rats receiving 200 mg/kg PA administration was better than those rats receiving 20 mg/kg BER administration, and almost compatible with the rats receiving 0.2 mg/kg TA administration.
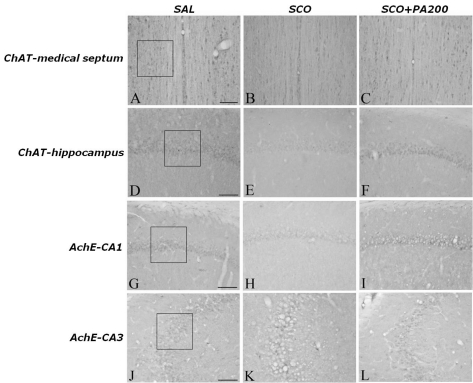 | Fig. 4Representative photographs showing the distribution of choline acetyltransferase- and acetylcholinesterase-reactive cells in the hippocampus of the SAL-ChAT-MS (A), SCO-ChAT-MS (B), SCO+PA200-ChAT-MS (C), SAL-ChAT-hippocampus (D) SCO-ChAT-hippocampus (E), SCO+PA200-ChAT-hippocampus (F), SAL-AchE-CA1 (G), SCO-AchE-CA1 (H), SCO+PA200-AchE-CA1 (I), SAL-AchE-CA1 (J), SCO-AchE-CA1 (K), and SCO+PA200-AchE-CA1 (L) groups. Sections were cut coronally at 30 µm. Scale bar indicates 50 µm.
|
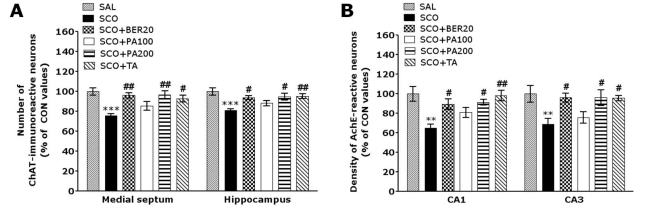 | Fig. 5Percentage (±SE) values of the mean number of choline acetyltransferase (ChAT)-stained neurons (A) and the density of acetylcholinesterase-stained neuron (B) in different hippocampal areas after the Morris water maze task. **p<0.01 and ***p<0.001 vs. the SAL group; #p<0.05 and ##p<0.01 vs. the SCO group.
|
Effect of PA and its main alkaloid compound, BER on hippocampal acetylcholinesterase
The density of AchE-immunopositive fibers in the CA1 and CA3 area of the rat hippocampus was significantly decreased in the SCO group as compared to the SAL group (p<0.01; Fig. 5B). The AchE-reactive neuronal activity in the CA1 and CA3 area of hippocampus due to SCO-induced memory impairment was significantly restored in the SCO+BER20 group (p<0.05) and the SCO+PA200 group (p<0.05), as compared to the SCO group. The effect of PA and BER on the density of AchE reactive neurons in hippocampus CA3 region was similar to that in the CA1 region. It also indicated that the density of AchE-reactive neurons in the hippocampus in rats receiving 200 mg/kg PA administration was better than those rats receiving 20 mg/kg BER administration, and almost compatible with the rats receiving 0.2 mg/kg TA administration.
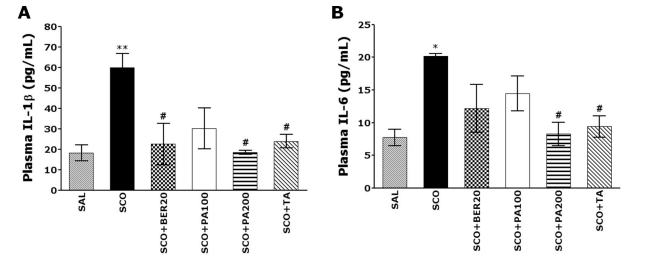
Effect of PA and its main alkaloid compound, BER on the IL-1β and IL-6 protein expression in the plasma
The ELISA analysis demonstrated that SCO administration for 14 days significantly increased the plasma IL-1β and IL-6 proteins expression in the rats by 229.07% and 161.39%, respectively. The expression of IL-1β protein in the plasma was markedly increased in the SCO group, as compared to the SAL group (p<0.01; Fig. 6A). The expression of IL-1β protein in the plasma due to SCO-induced memory impairment was significantly restored in the SCO+BER20 group (p<0.05) and the SCO+PA200 group (p<0.05), as compared to SCO group. The expression of IL-6 protein in the plasma was also markedly increased in the SCO group, as compared to the SAL group (p<0.05; Fig. 6B). The expression of IL-6 protein in the plasma due to SCO-induced memory impairment was significantly restored in the SCO+PA200 group (p<0.05), as compared to SCO group. It also indicated that the expression of IL-1β and IL-6 proteins in the plasma in rats receiving 200 mg/kg PA administration was better than those rats receiving 20 mg/kg BER administration, and almost compatible with the rats receiving 0.2 mg/kg TA administration.
Effect of PA and its main alkaloid compound, BER on BDNF, CREB, IL-1β, TNF-α and COX-2 mRNA expression levels in the hippocampus
The effect of PA or BER administration on BDNF, CREB, IL-1β, TNF-α and COX-2 mRNA expression levels in the rats with SCO-induced hippocampus lesions was investigated using RT-PCR analysis (Fig. 7). Hippocampal BDNF and CREB mRNA expression in the SCO group were significantly decreased as compared to that of the SAL group (p<0.01). The decreased expression of BDNF and CREB mRNA in the SCO group was significantly restored in the SCO+BER20 group (p<0.05) and the SCO+PA200 group (p<0.01). It also indicated that the expression of BDNF and CREB mRNA in the hippocampus in rats receiving 200 mg/kg PA administration was better than those rats receiving 20 mg/kg BER administration, and almost compatible with the rats receiving 0.2 mg/kg TA administration. Hippocampal IL-1β and TNF-α mRNA expression in the SCO group was significantly increased as compared to that of the SAL group (p<0.05 and p<0.01, respectively). The increased expression of IL-1β mRNA in the SCO group was significantly restored in the SCO+PA200 group (p<0.05). The increased expression of TNF-α mRNA in the SCO group was also significantly restored in the SCO+PA200 group (p<0.01). It also indicated that the expression of IL-1β and TNF-α mRNA in the hippocampus in rats receiving 200 mg/kg PA administration was better than those rats receiving 20 mg/kg BER administration, and almost compatible with the rats receiving 0.2 mg/kg TA administration. Hippocampal COX-2 mRNA expression in the SCO group were also significantly increased as compared to that of the SAL group (p<0.01). The increased expression of COX-2 mRNA in the SCO group was significantly restored in the SCO+PA100 group (p<0.05) and the SCO+PA200 group (p<0.05). It also indicated that the expression of COX-2 mRNA in the hippocampus in rats receiving 200 mg/kg PA administration was almost compatible with the rats receiving 0.2 mg/kg TA administration.
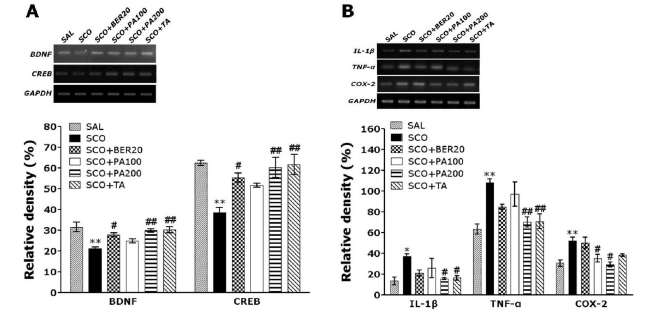 | Fig. 7PCR bands and their relative intensities of brain-derived neurotrophic factor (BDNF) and cAMP-response element-binding protein (CREB) mRNA (A), and PCR bands and their relative intensities of interleukin-1 beta (IL-1β), tumour necrosis factor (TNF)-α, and cyclooxygenase-2 (COX-2) mRNA (B) in the hippocampus of scopolamine-induced cognitive impairment rats. *p<0.05 and **p<0.01 vs. the SAL group; #p<0.05 and ##p<0.01 vs. the SCO group.
|
Go to :
DISCUSSION
PA is well known to have multiple pharmacological activities and has been used for treating various cognitive disorders and psychosomatic diseases such as stress [20]. Although typical prescriptions including PA are known to be clinically effective in alleviating memory-related symptoms, the underlying mechanisms of PA in those effects have not been investigated. A major component of PA is BER, which could potentially possess anti-AD effects [21]. Recently, some studies demonstrated that BER could inhibit the expression of some inflammatory factors through the nuclear factor-κB (NF-κB) signaling pathway [22]. Also, BER can penetrate the blood-brain barrier to reach the striatum, cortex, and hippocampus, and thus, BER might act directly in brain nuclei to produce neuroprotective effects [11]. However, our results are not consistent with the previous studies showing that BER could inhibit memory impairment and expression of the inflammatory factor interleukin-1β by increasing microglial activation [13]. The difference between our results and those of previous studies may result from differing effects of BER on immune and inflammatory factor expression in different animal modes of Alzheimer's disease.
The present results demonstrated that repeated SCO-induced dementia produced severe deficits in performance on tests of cognitive functioning as well as corresponding signs of neurodegeneration in the brain, including decreased ChAT and AchE activities in the hippocampus. Our results showed that administration of PA and BER significantly improved learning and memory on the PAT and it decreased the escape latency and increased the number of platform crossings in the MWM test in a rat model of AD. It also produced increased ChAT and AchE immunoreactivity in septo-hippocampal cholinergic neurons areas and increased BDNF and CREB mRNA expression in the hippocampus associated with the SCO-induced memory impairment in male rats. PA and BER also inhibited the increase in proinflammatory cytokines in the plasma and hippocampus of rats with SCO-induced deficits.
Our results suggest that administration of 200 mg/kg PA produced a significantly higher inhibitory effect on activity than did 20 mg/kg BER. It is known that PA contains palmatine and coptisine as well as BER, and previous reports have shown that palmatine and coptisine regulated behavioral activity and had CNS effects. Thus, other alkaloids may also affect SCO-induced memory deficits and the cholinergic system.
Chronic administration of SCO in animals transiently produces several deficits in learning acquisition and short-term memory that are considered to be similar to the cholinergic deficits in AD or senile CNS dysfunction [3]. The well-replicated amnesic effect of SCO has been interpreted as a principal consequence of the blockade of post-synaptic muscarinic M1 transmission, leading to disruption in the functioning of the hippocampus in working memory. Thus, the SCO-induced amnesic model has been widely used as a pharmacological model of memory dysfunction in screening for agents with potential cognition-enhancing effects [4].
This study showed that SCO significantly shortened the step-through latency of the retention trial in the PAT, demonstrating that the central cholinergic neuronal system plays an important role in learning acquisition and synaptic plasticity. PA and BER prolonged the step-through latency of the passive avoidance response that had been shortened by SCO. These results suggest that the anti-amnesic effect of PA and BER in SCO-induced memory impairment may be mediated by the cholinergic nervous system.
To confirm the effects of PA and BER on other types of memory, we performed the MWM test to evaluate spatial learning. The MWM test is a hippocampus-dependent memory task that is frequently used for examining cognitive deficits and demonstrating permanent spatial learning ability and reference memory in rodents [23]. Animals encode spatial working information during the learning step, which then serves to guide future memory retrieval. The current finding that the memory deficits demonstrated by the SCO-impaired rats in the MWM task were more pronounced than were those in the SAL group were consistent with a previous study [24]. During the trial sessions, the PA and BER groups showed significantly reduced escape latency, enhanced cognitive performance, and amelioration of the memory deficits associated with SCO. In terms of average swim speeds and rest times, both indices of motor functioning, the SCO group did not differ significantly from the PA-treated and BER-treated groups, suggesting that motor impairment was not the primary cause of the poor performance of the SCO group in the MWM test. Thus, it is evident that PA and BER administration improved MWM performance by directly enhancing spatial working memory. Scores on the escape and the spatial probe tests of the MWM test are considered to primarily reflect long-term spatial memory ability. In this study, PA and BER treatment shortened the escape latency without affecting swimming velocity and extended the time spent swimming in the place where the platform was previously located. In the probe test, administration of PA and BER lowered the escape latency to levels comparable to those demonstrated by the control group and significantly increased the number of times that the rats crossed the platform location [25].
Our results showed that chronic administration of SCO also produced severe deficits in cholinergic neuron dysfunction as well as decreased ChAT and AchE activities in the hippocampus, indicating neurodegeneration in the brain. Impairment in the hippocampus generally results in diminished spatial learning ability [23,26]. It seems likely that the central cholinergic neuronal system plays an important role in learning acquisition and synaptic plasticity [27]. The expression and activation of AchE and ChAT regulate the dynamic concentration of ACh in the cholinergic synapses in the brain [28]. PA and BER improved cholinergic neurons in the hippocampus and continuously induced increases in ChAT and AchE activities, which eventually resulted in recovery of the entire cholinergic circulation pathway [29]. According to the cholinergic hypothesis, memory impairment in patients with senile dementia is due to selective and irreversible deficits in cholinergic function or alternation in hippocampal functioning in the brain [29]. Thus, cholinesterase inhibitors and ChAT activators may compensate for reduced the ACh levels in brains with AD disease. Previous biochemical and behavioral evidence indicated that central cholinergic transmission declines as a result of SCO administration [30] and the type of dementia associated with AD [31]. This type of decline has been closely associated with cognitive disturbances. It seems likely that the observed amelioration in the deficits in spatial learning capability demonstrated in the PA-treated and BER-treated rats were associated with the increased release of ACh. Thus, we propose that the beneficial effects of PA and its major alkaloid compound, BER, to ameliorate memory impairments could be related to increase central cholinergic functioning.
On the other hand, new findings concerning the activation of BDNF and CREB as novel therapeutics have been reported through several basic and clinical studies [32]. The intentional expression of BDNF and CREB proteins in the brain provides a novel treatment strategy for healing memory deficits [33]. Recent experimental evidence strongly supports the role of BDNF in the modulation of synaptic function and plasticity in CNS for learning and memory processes, besides its actions on neuronal cell survival and prevention of neurodegeneration [33]. Also, there is sufficient evidence that the CREB regulates the expression of genes involved in neuroplasticity, cell survival, and cognition [34]. The phosphorylated CREBs are able to bind to cAMP response elements of the target genes considered to be involved in long-term memory formation [35]. Thus, BDNF transcription, regulated by CREB, may also be a critical player in the adaptive neuronal responses underlying learning and memory function [36]. Thus, a previous study suggested a close correlation between reduced expression of BDNF and CREB in the hippocampus and cognitive impairment [37]. SCO-induced memory deficits induced significant reductions of the BDNF and CREB mRNA expression in the hippocampus and poor performance in the hippocampus-dependent tasks [38,39]. We thus proposed that administration of PA and BER significantly prevented the reduction of the BDNF and CREB mRNA expression induced by SCO. These findings suggested that the beneficial effects of PA and BER reduces SCO-induced memory and learning deficits by the increase in BDNF expression via the CREB signaling pathway and could be related to increase in neuronal functioning.
At the same time, many studies have suggested that amyloid β-peptide (25-35)-induced or SCO-induced IL-1β and TNF-α expression is upregulated in AD [40] and that these cytokines may play a role in several events in the pathological cascade of AD [41]. These results suggest that inflammatory reactions may be related to the pathogenesis of degenerative changes and cognitive impairment [42]. In the present study, SCO administration caused an increase in the proinflammatory cytokines IL-1β and TNF-α as well as learning and memory deficits. PA and BER inhibited the increased activation of proinflammatory cytokines such as IL-1β and TNF-α in SCO-treated rats.
Many studies have suggested that the inducible gene encoding COX-2 is a key element that modulates the generation of proinflammatory mediators including various prostagladins [43]. It is known that the expression of COX-2 and the synthesis of prostaglandin E2 (PGE2), one of its products, are increased in the hippocampus of AD patients, which may be related to the pathogenesis of the degenerative changes and cognitive impairments [44]. These results suggest that inflammatory responses to SCO injection significantly stimulated the expression of COX-2 mRNA in the hippocampus through modulating the NF-κB pathway [45]. The increase in COX-2 expression by NF-κB activation can accelerate inflammatory responses and subsequently contribute to learning and memory deficits. Thus, treatment with long-lasting COX-2 inhibitors during the initial stages of AD, before clinical symptoms of dementia appear, may suppress inflammatory responses and the synthesis of pro-inflammatory mediators in the brain [46]. In the present study, PA significantly decreased SCO-stimulated behavioral changes and memory disturbances through inhibition of COX-2 mRNA expression. Thus, we proposed that the anti-inflammatory effects of PA and BER significantly reversed impairment in spatial working memory.
The present study showed that repeated SCO-induced dementia produced learning and memory deficits as well as associated degeneration of cholinergic neurons, impairment of neuronal function and inflammatory reactions in rats, as evidenced by their performances on the PAT and MWM tests and gene expression analysis in a rat model of AD disease. However, administration of PA and BER attenuated these SCO-induced dementia effects, as indicated by improved cognitive functioning during behavioral tests and increases in the density of cholinergic neurons. Thus, PA and its major alkaloid compound, BER may be useful agents in preventing cholinergic dysfunction, neuronal impairment, and anti-inflammatory effects such as those observed in AD.
Go to :
 XML Download
XML Download