

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
In addition to oxidized forms of cholesterol, multiple infectious agents including Chlamydia pneumoniae, Mycoplasma pneumoniae, Porphymonas gingivalis, Staphylococcus species, and Streptococcus species are found in the atherosclerotic plaque [1,2]. The pathogens comprise pathogen-associated molecular patterns (PAMPs) that are recognized by the toll-like receptors (TLRs). TLRs are type 1 transmembrane proteins with an ectodomain consisting of leucine rich repeats required for recognition of PAMPs, a transmembrane domain that determines cellular localization, and an intracellular toll interleukin 1 receptor (TIR) domain needed for downstream signaling [3]. Expression and function of TLRs are associated with atherosclerosis, which is caused by inflammation. For instance, mice deficient in TLR4 and TLR2 showed a 55% decrease in atherosclerotic lesions, while a 65% decrease in macrophage infiltration was observed in ApoE-/- mice deficient in TLR4 [4,5]. TLR6 forms a heterodimer with TLR2 and TLR4 [6,7]. Coexpression of TLR2 and TLR6 at the cell surface is crucial for recognition of diacylated lipopeptide from mycoplasma and peptidoglycan and subsequent cellular activation [8]. TLR6 activation induces production of tumor necrosis factor (TNF)-α in macrophages [9]. However, signaling molecules associated with TLR6 are clarified.
Chemokines and cytokines play important roles in atherosclerosis by promoting recruitment and migration of inflammatory cells into the atherosclerotic lesion and by inducing activation of endothelial cells and leukocyte subsets. The activated cells, in turn, release more cytokines and chemokines, leading to enhanced inflammatory responses in atherosclerotic lesions [10-12]. Chemokine (C-C motif) ligand 2 (CCL2), monocyte chemotactic protein-1 (MCP-1), is critical for recruitment of monocytes to atherosclerotic plaques and its expression has been linked to atherosclerosis [13]. Deletion of the CCL2 gene or the gene for its receptor resulted in reduced development of atherosclerotic plaques [14]. The decrease in lesion size in the absence of TLRs was associated with a decrease in peripheral levels of CCL2 [4]. In addition to CCL2, expression of interleukin-1 (IL-1), a prototypic proinflammatory cytokine that stimulates both local and systemic inflammatory responses to multiple infectious agents, is also elevated in macrophages of human atherosclerotic plaques [15]. Genetic deficiency and inhibition of IL-1β and its receptor IL-1R1 resulted in a decrease in atherosclerotic plaque formation in animal models of atherosclerosis [16-18]. IL-1 protein is considered to induce production of cytokines and chemokines and increases expression of adhesion molecules on endothelial cells, thus enhancing the recruitment of inflammatory cells [16-18]. Therefore, due to their close association with atherosclerosis, understanding the regulation of CCL2 and IL-1 expression is important.
Although TLR6 is believed to participate in inflammatory responses through forming heterodimers with TLR2 and TLR4 [6,7], the proinflammatory property of TLR6 itself is poorly characterized. To test the hypothesis that TLR6 is able to activate signaling pathways contributing to or leading to inflammation, we evaluated the ability of the TLR6 agonist FSL-1 (Pam2CGDPKHPKSF), a synthetic lipoprotein derived from Mycoplasma salivarium, to induce CCL2 and IL-1β expression in macrophages. In addition, we determined signaling molecules downstream of TLR6 that were involved in FSL-1-mediated upregulation of CCL2 and IL-1β.
METHODS
Cells
THP-1 cells were cultured in RPMI medium 1640 supplemented with 10% fetal bovine serum (FBS) in a humidified atmosphere of 5% CO2 in the presence of penicillin (50 units/ml) and streptomycin (50 µg/ml). THP-1 cells were passaged every 2~3 days in order to maintain between 1,000 to 1,000,000 cells per ml in the culture medium. THP-1 cells in passages between 7 and 10 were used for experiments.
Reagents
FSL-1 and oxidized 1-palmitoyl-2-arachidonosyl-sn-phosphatidylcholine (OxPAPC) were purchased from Invivogen (San Diego, CA, USA). LY294002, RO318220, GF109203X, and SP600125 were purchased from Sigma-Aldrich (St. Louis, MO, USA). U0126, SB202190, and Akt inhibitor IV (Akti IV) were purchased from Cell Signaling Technology (Danvers, MA, USA). Anti-phospho-ERK antibody was purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Anti-phospho-p38 MAPK antibody was purchased from R&D systems (Minneapolis, MN, USA). Anti-phospho-Akt antibody and anti phospho-JNK antibody were purchased from Cell Signaling Technology (Beverly, MA, USA). Anti-α-tubulin antibody was purchased from Calbiochem.
Enzyme-linked immunosorbent assay (ELISA)
The amount of CCL2 and IL-1β released from THP-1 cells into the culture medium was determined using commercially available ELISA kits according to the manufacturer's instructions (R&D Systems, Minneapolis). Recombinant standards of CCL2 or IL-1β provided in the kit and the isolated culture medium were added to a plate pre-coated with a monoclonal antibody against the chemokine. After incubation for 2 h, the plate was washed and incubated with an enzyme-linked polyclonal antibody specific for CCL2 or IL-1β. After several washes, the substrate solution was added, and the color intensity was measured. A standard curve was used for determination of the amount of CCL2 or IL-1β present in the samples. Data are expressed as average±standard deviation of triplicates experiments.
Reverse transcription (RT)-polymerase chain reaction (PCR)
RT-PCR and real-time PCR were performed as previously described [19]. In brief, total RNAs isolated were reverse-transcribed for one hour at 42℃ with Moloney Murine Leukemia Virus reverse transcriptase, followed by PCR analysis using primers. The cDNA was denatured at 90℃ for 5 min followed by 25 cycles of PCR (95℃ for 30 sec, 55℃ for 30 sec, 72℃ for 30 sec). The primers for CCL2 were 5-TCTGTGCCTGCTGCTCATAG-3 (forward) and 5-CAGATCTCCTTGGCCACAAT-3 (reverse), and the primers for IL-1β were 5-GGACAAGGGAGGAAGATGC-3 (forward) and 5-TCTTTCAACACGCAGGACAG-3 (reverse). Transcripts of glyceraldehydes-3-phosphate dehydrogenase (GAPDH) were amplified as an internal control. Primers for GAPDH were 5-GAGTCAACGGATTTGGTCCT-3 (forward) and 5-TGTGGTCATGAGTCCTTCCA-3 (reverse). The intensities of CCL2 and IL-1β bands were normalized relatively to that of GAPDH which was not changed by each treatment. Error bars represents the standard deviation of triplicates experiments.
Western blot analysis
THP-1 cells were lysed with lysis buffer (1% SDS, 1 mM NaVO3, 10 mM Tris-HCl, pH 7.4) containing protease inhibitors, and supernatants were isolated after centrifugation (15,000×g, for 5 min, at 4℃). Cell lysates containing an equal amount of protein were separated by 12% SDS-PAGE and transferred to polyvinylidene fluoride membranes. After blocking for one hour in 5% skim milk in 0.1% Tween 20/TBS, the membranes were incubated at 4℃ with appropriate secondary antibody diluted in blocking solution overnight. After washing three times with 0.1% Tween 20/TBS for 10 min each, the membranes were incubated for one hour at room temperature with horseradish peroxidase-conjugated secondary antibodies diluted in blocking solution (1:5,000). After washing three times with washing buffer for 10 min each, bands were detected with chemiluminescent reagents.
RESULTS
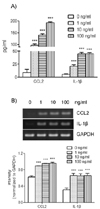
Upregulation of CCL2 and IL-1β in response to FSL-1 at the messenger and protein levels
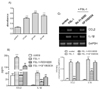
In order to examine the effects of TLR-6 activation on expression of CCL2 and IL-1β in THP-1 cells, we performed concentration and time course experiments using FSL-1. Results of ELISA analyses revealed that THP-1 cells significant increase in the presence of FSL-1. The amount of CCL2 secreted increased by 7.2-, 10.3-, and 14.3-fold in the presence of 1, 10 and 100 ng/ml of FSL-1, respectively, in comparison with control cells cultured in the absence of FSL-1 (Fig 1A). The amount of IL-1β secreted was increased by 3.9-, 8.3-, and 8.2-fold in the presence of 1, 10, and 100 ng/ml of FSL-1, respectively, compared with control cells. We also performed RT-PCR in order to determine whether FSL-1 had an influence on expression of CCL2 and IL-1β at the mRNA level. CCL2 and IL-1β gene transcripts were barely detected in THP-1 cells in the absence of FSL-1, whereas their transcripts were induced in the presence of 1, 10, and 100 ng/ml of FSL-1 (Fig. 1B). FSL-1-enhanced secretion of CCL2 and IL-1β was observed as early as 3 h post-treatment, and appeared to reach a plateau at 6 h post-treatment, and was sustained up to 12 h post-treatment with FSL-1 (Fig. 2A). FSL-1-induced transcription of CCL2 was observed as early as 3 h post-treatment transcripts, and the induction became more evident 6 h post-treatment and thereafter. FSL-1 induced transcription of IL-1β as early as 3 h post-treatment transcript and persisted up to 12 h post-treatment with FSL-1 (Fig. 2B).
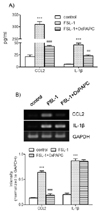
Participation of TLR2/4 in FSL-1-induced expression of CCL2 and IL-1β
Since TLR6 co-operates with TLR2/4 via heterodimerization to induce expression of proinflammatory genes [6-8], we investigated the question of whether TLR2/4 played a role in FSL-1-mediated expression of CCL2 and IL-1β using OxPAPC, the TLR2/4 inhibitor. OxPAPC had a profound influence on secretion of CCL2 and IL-1β (Fig. 3A). Secretion of CCL2, which increased by 10.6-fold in the presence of FSL-1, was reduced to 1.9-fold and secretion of IL-1β, which increased by 8.9-fold in response to FSL-1, was attenuated to 4.5-fold in the presence of OxPAPC in comparison with control cells. The TLR2/4 inhibitor affected transcription of CCL2 but not of IL-1β. In particular, upregulation of CC2 transcripts induced by FSL-1 was abrogated in the presence of the inhibitor (Fig. 3B).
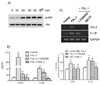
Roles of PI3K-Akt in FSL-1-induced expression of CCL2 and IL-1β
To investigate the effect of FSL-1 in the activity of Akt, we performed western blot analysis in order to examine phosphorylation of Akt. Treatment with FSL-1 resulted in enhanced phosphorylation of Akt. Maximum Akt phosphorylation occurred 20 min post-treatment with FSL-1, and was sustained up to 40 min post-treatment (Fig. 4A). We investigated the roles of Akt in FSL-1-mediated expression of CCL2 and IL-1β using inhibitors of LY294002 and Akti IV. LY294002 inhibited phosphoinositide 3-kinase (PI3K), leading to activation of Akt kinase. Both inhibitors had a significant effect on expression of CCL2 at the messenger and protein levels. Secretion of CCL2 increased by 10.2-fold in response to FSL-1, which was almost completely blocked in the presence of LY294002 and Akti IV (Fig. 4B). In addition, both inhibitors abrogated FSL-1-mediated transcription of CCL2 (Fig. 4C). Treatment with LY294002 and Akti IV also resulted in significantly attenuated secretion of IL-1β (Fig. 4C). Secretion of IL-1β increased by 8.1-fold in response to FSL-1. The increase was attenuated to 3.5-fold and was abrogated in the presence of LY294002 and Akti IV, respectively (Fig. 4B). In addition, treatment with Akti IV, but not LY294002, resulted in abrogated IL-1β gene transcription induced by FSL-1.
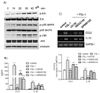
Major roles of the ERK1/2 pathway in FSL-1-induced expression of CCL2 and IL-1β
We attempted to determine whether FSL-1 activated mitogen-activated protein kinases (MAPKs) by detection phosphorylated forms of extracellular signal-regulated kinase (ERK), p38 MAPK, and c-jun N-terminal kinase (JNK) on western blots (Fig. 5A). Treatment with FSL-1 resulted in enhanced phosphorylation of ERK, which was observed 20 min post-treatment and reached maximum 30 min post-treatment with FSL-1. FSL-1 also affected phosphorylation of p38 MAPK and JNK; maximum phosphorylation of the kinases occurred 20 min post-treatment with FSL-1 and was sustained up to 40 min post-treatment with FSL-1. We used inhibitors of SB202190 (a p38 MAPK inhibitor), SP600125 (a JNK inhibitor), and U0126 (an ERK inhibitor) in order to assess the roles of MAPKs in FSL-1-induced upregulation of CCL2 and IL-1β. The three MAPK inhibitors had a significant influence on secretion of CCL2 and IL-1β in a similar pattern. Secretion of CCL2 increased by 9.7-fold in response to treatment with FSL-1, which was abrogated by the treatment with U0126 and reduced to 5.2- and 2.6-fold in the presence of SB202190 and SP600125, respectively (Fig. 5B). Secretion of IL-1β increased by 9.7-fold in response to FSL-1. The increase was lowered to 1.5-, 5.3-, and 4.0-fold in the presence of U0126, SB202190, and SP600125, respectively. The MAPK inhibitors also influenced CCL2 and IL-1β expression at the mRNA level in a pattern similar to that observed with CCL2 and IL-1β secretion (Fig. 5C). Gene transcription of CCL2 and IL-1β induced in the presence of FSL-1 was abolished by treatment with U0126 and attenuated by treatment with SB202190 and SP600125.
Involvement of PKC in FSL-1-induced expression of CCL2 and IL-1β
Examining the effects of the TLR6 ligand on protein kinase C (PKC), we found that FSL-1 enhanced the activity of PKC (Fig. 6A). Increased PKC activity was observed as early as 5 min post-treatment and remained enhanced up to 30 min post-treatment with FSL-1. To assess the role of PKC in FSL-1-mediated upregulation of CCL2 and IL-1β, we used two PKC inhibitors, GF109203X and RO318220. Both inhibitors affected secretion of FSL-1-mediated CCL2 and IL-1β in a similar pattern. Secretion of CCL2 increased by 9.4-fold in response to the treatment with FSL-1, which was lowered to 1.5- and 5.7-fold in the presence of RO318220 and GF109203X, respectively (Fig. 6B). Secretion of IL-1β was enhanced by 8.1-fold in response to treatment with FSL-1, and was attenuated to 2.1- and 4.9-fold in the presence of RO318220 and GF109203X, respectively. Inhibition of PKC also influenced expression of CCL2 and IL-1β at the mRNA level. FSL-1-mediated CCL2 and IL-1β gene transcription was profoundly inhibited by treatment with RO318220 not by GF109203X (Fig. 6C).
DISCUSSION
In the current study, we demonstrated that treatment with FSL-1, a bacterial-derived TLR6 agonist, resulted in upregulated expression of CCL2 and IL-1β both at the mRNA and protein levels in human macrophage THP-1 cells. This finding is consistent with the fact that TLRs cause upregulation of inflammatory gene transcription upon recognition of their ligands [3,20]. We also attempted to identify cellular factors contributing to FSL-1-mediated expression of CCL2 and IL-1β. TLRs can form homodimers or heterodimers. TLR2 forms a heterodimer with TLR-1 or -6 and TLR4 forms a heterodimer with TLR6. Co-expression of them is necessary for gene expression [6-8]. Therefore, we attempted to investigate whether TLR2/4 played roles in FSL-1-induced expression of CCL2 and IL-1β. Treatment with OxPAPC markedly resulted in markedly blocked FSL-1-mediated secretion of CCL2 and abrogated FSL-1-induced CCL2 gene transcription, indicating requirement of TLR2/4 activity. However, OxPAPC did not influence FSL-1-induced IL-1β gene transcription, yet treatment with the inhibitor resulted reduced FSL-1-mediated IL-1β secretion by approximately 50%. The differential regulation of CCL2 and IL-1β expression in the presence of OxPAPC can be explained in conjunction with TLR2/4. FSL-1 is able to activate TLR6 via TLR2/4-dependent and -independent ways. FSL-1 induces CCL2 via TLR2/4-dependent mechanism while TLR2/4-independent signaling pathway seems to play a major role in transcription of IL-1β in response to FSL-1.
The PI3K pathway regulates TLR signaling, leading to either positive or negative effects on signaling, depending on cell type and stimulus [21,22]. We found that FSL-1 enhanced phosphorylation of Akt, suggesting that the TLR6 ligand activated the PI3K pathway, as PI3K activation leads to phosphorylation of the Akt kinase [23,24]. We attempted to determine whether Akt and PI3K, the Akt activator, were involved in expression of CCL2 and IL-1β. Inhibition of Akt and PI3K resulted in complete blockade of FSL-1-mediated secretion of CCL2 as well as CCL2 gene transcription. Inhibition of Akt resulted in blockade of FSL-1-mediated expression of IL-1β at the protein and the mRNA levels, while inhibition of PI3K reduced resulted in FSL-1-mediated secretion of IL-1β by approximately 35% without noticeable decrease of its gene transcription. These findings indicate that both PI3K and Akt are necessary for FSL-1-mediated CCL2 expression, whereas Akt is sufficient for IL-1β expression in response to FSL-1. The failure of LY294002 to blocked transcription of IL-1β might be explained in relation to the mode of action of LY294002 as well as to pattern of IL-1β expression. LY294002 is a reversible PI3Ks inhibitor [25], indicating that inhibition of PI3Ks depends upon relative activity between LY294002 and PI3K activation by treatment with FSL-1. In addition, transcription of IL-1β occurred fast after FSL-1 treatment. It may be possible that signaling pathways activated FSL-1 suppassed inhibitory activity of LY294002 for duration such that IL-1β, but not CCL2, gene transcription was induced.
MAPKs, the serine/threonine-specific protein kinases that respond to extracellular stimuli and regulate various cellular activities, mediate chemokine production in response to activation of TLR-2, -4, and -9 [26-28]. We found that treatment with FSL-1 resulted in elevated phosphorylation of ERK, p38 MAPK, and JNK, indicating that FSL-1 activated the three MAPKs. We investigated the question of whether MAPKs played roles in FSL-1-mediated expression of the chemokine and cytokine using specific pharmacological inhibitors of the kinases. Inhibition of ERK resulted in abrogation of secretion of CCL2 and IL-1β and transcription of the two genes. Inhibition of p38 MAPK and JNK also resulted in significantly attenuated FSL-1-mediated expression of CCL2 and IL-1β primarily at the protein level. Our results indicate that ERK activity is required for FSL-1 mediated expression of CCL2 and IL-1β at the messenger and protein levels and that p38 MAPK and JNK are necessary for maximum expression of CCL2 and IL-1β in response to FSL-1.
THP-1 cells express PKC-βI, -βII, -λ, -η, and -θ subtypes [29]. Pharmacological inhibition of PKC, or its depletion by long-term treatment with phorbol esters, results in decreased secretion of TLR-2 and -4-stimulated cytokines and chemokines [27,30,31]. These reports suggest the possible involvement of PKC in FSL-1-mediated gene expression. We investigated the effects of FSL-1 on PKC activity and found that FSL-1 enhanced PKC activity. Therefore, we attempted to determine whether PKC played roles in FSL-1-mediated expression of CCL2 and IL-1β using PKC inhibitors. FSL-1-mediated expression of CCL2 and IL-1β was profoundly inhibited by RO318220 in comparison with GF109203X. The two inhibitors are bisindolylmaleimide derivatives of staurosporine and inhibit mixed isoforms of PKC [32]. In addition to PKC, RO318220 inhibits Akt, c-Raf, MAPKK-1, and p42 MAPK, however, GF109203X does not [32]. In the current study we demonstrated that activities of Akt and p44/42 MAPK were required for FSL-1-mediated expression of CCL2 and IL-1β. It is possible that the additional inhibition of kinase activity including Akt and p44/42 MAPK by RO318220 contributed to the difference between the two inhibitors in their effects on expression of CCL2 and IL-1β. Our data suggest involvement of PKC activity in FSL-1-induced gene transcription of IL-1β and CCL2. More investigation is necessary to clarify roles of PKCs in the FSL-1 action.
Our results indicate that FSL-1, a TLR6 ligand, upregulates CCL2 or IL-1β expression by activating PI3K-Akt pathway, MAPKs, and PKC in a TLR2/4-dependent and -independent mechanisms. Our results suggest that TLR6 activation contributes to inflammatory responses by upregulation of inflammatory chemokines and cytokines, including CCL2 and IL-1β, via activation of multiple signaling pathways in macrophages.

 XML Download
XML Download