

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Voltage-dependent ion channels are major contributors to decide physiological functions and the generation of pathophysiological conditions in mammalian excitable cells, such as neurons and cardiac myocytes. Therefore, agonists and antagonists of ion channels are widely used for biological and therapeutical approaches in researches and clinic applications [1-5]. In nature, a number of toxins acquired from insects, snakes, and fish play roles to block and modulate the permeability of ion channels expressed in cellular membranes. For example, Na+ channels are blocked by tetrodotoxin from pufferfish and mu-conotoxin from Conusstriatus, while they are opened by versutoxin from spiders [6-8]. Because the pharmacological modification of gating properties of ion channels is most simple but powerful for testing their functions in neurons, the discovery of new toxins also leads biological researchers to find unknown physiological functions of cells that are correlated with specific ion channels in mammalian systems. In addition, most ion channels have several subunits exhibiting distinctive kinetics and permeability, indicating that the classification of specific toxins for each ion channel subunit is also important to define their delicate functions [9-12].
As they can determine cardiac rhythm and neuronal transmission, voltage-dependent K+ channels (Kv channels) are most critical channels working in both cardiac and nervous systems [13-16]. In electrophysiological experiments, major K+ outward currents through Kv channels can be recorded by using voltage-clamp methods and classified into two patterns, transient or rapidly inactivating currents - known as A-type (IA) - and sustained or not inactivating currents [13,16]. In hippocampal CA1 neurons of mammalian brains, Kv4.2 and Kv4.1 subunits of Kv channels are dominant for transient currents, while Kv2.1 contributes to sustained currents [13,17]. Previous papers have reported that these K+ subunits are sensitive to several natural toxins and chemicals [16,18-21]. However, specific agents working for each subunit are still obscure. For example, hanatoxins have been reported to block Kv4.2 as well as Kv2.1 channel subunits expressed in Xenopus laevis oocytes, indicating that these toxins are not specific for one of several subunits [18]. Recently, heteropodatoxins (HpTXs) isolated from a spider, Heteropoda venatoria, are known as more specific blockers of IA by blocking Kv4 families but not Kv1.4 subunits. In particular, HpTX2 has dose- and voltage-dependent blockade effects on IA via binding the S3-S4 linker region of Kv4.2 channels [19]. These previous results are not sufficient to define and specify the effects of HpTX2 on IA channels, because the blocking effects were tested only on channels expressed experimentally in Xenopus oocytes or cardiac cells. In addition, subunits of voltage-dependent K+ channels in native mammalian neurons have homologue structures with six membrane spanning portions and share auxiliary subunits, insisting that the effects of HpTX2 on K+ currents must be demonstrated in neurons [22]. It is also possible that, because of the hydrophilic property of the toxin and the location of binding sites, HpTX2 might have effects on other K+ subunits according to application protocols. In the present study, we attempted to test effects of HpTX2 on both transient and sustained K+ outward currents in CA1 neurons of cultured organotypic hippocampal slices. In nucleated-patch experiments, extracellular HpTX2 only reduced transient currents. However, when HpTX2 was added to internal pipette solution, sustained but not transient currents showed a significant reduction. Although we did not discover the binding mechanism in this study, these results clearly indicate that HpTX2 effects to modulate or block K+ outward currents may be dependent on the location of binding sites of each subunit.
METHODS
Tissue preparation
Organotypic hippocampal slices (350 µm thick) were prepared from postnatal day 7~8 Sprague-Dawley rats. After preparation of hippocampal slices in a cutting solution containing (in mM): 10 MgCl2, 25 Glucose, 20 Hepes in Gey's balanced salt solution (Sigma), slices were transferred to an incubator gassed with 95% O2 and 5% CO2 at 35℃, and cultured for 5~6 days before performance of electrophysiological measurements. More detailed protocols for tissue preparation are available in our previous paper [16]. The Animal Care and Use Committee of Jeju National University approved all animal protocols.
Electrophysiology
For patch-clamp recordings from organotypic slices, slices were transferred to a submerged recording chamber with continuous flow of ACSF containing (in mM): 125 NaCl, 2.5 KCl, 25 NaHCO3, 1.25 NaH2PO4, 25 Glucose, 2 CaCl2, 1 MgCl2. 5 µM 2-chloroadenosine, and 5 µM bicuculine were added in some recordings. TTX (1 µM) was added for recording voltage-dependent K+ currents immediately after pulling nucleated-patches. Normal patch electrodes (4~6 MΩ) were filled with (in mM): 20 KCl, 125 K-gluconate, 10 HEPES, 4 NaCl, 0.5 EGTA, 4 ATP, 0.3 TrisGTP, and 10 Phosphocreatin. In some experiments, patch electrodes were filled with Cs-gluconate internal solution, including (in mM): 100 Cs-gluconate, 5 MgCl2, 0.6 EGTA, 8 NaCl, 40 HEPES, 2 NaATP, and 0.3 TrisGTP. pH and osmolarity were adjusted to 7.2~7.3 and 280~300 mOsm, respectively, in all experiments. K+ outward current recordings using a nucleated-patch were performed at 31~32℃ and low-pass filtered at 5 kHz and digitized at 10 kHz by an HEKA 9/2, which was controlled by software PULSE (HEKA). Command pulse generation and data acquisition were performed using PULSE, and IGOR Pro (Wavemetrics) and Excel (Microsoft) software were used for further data and statistical analysis. Unpaired t-tests were used in performance of statistical analysis, and significance was set to p<0.05 or 0.01.
K+ outward currents recordings
Recording voltage-gated K+ currents was made in voltage-clamp mode of nucleated-patches (18.35±0.5 pF, n=37). HpTX2 recombinant (Sigma-Aldrich, USA) was added to either external ACSF or internal pipette solution to test blocking effects on K+ outward currents, and 4-AP was added to internal pipette solution. In this study, to clarify the effects of drugs added intracellularly, all nucleated-patches were pulled within 2 min after making a whole-cell. Current ensemble averages were constructed from three individual sweeps. Leakage and capacitive currents were subtracted digitally using either a P/6 protocol or null traces. The transient current was isolated from the sustained current using 200 ms pre-pulse step to -20 mV for inactivation of transient channels. More detailed protocols for isolation of each pure current can be found in our previous paper [16].
RESULTS
Kv4.2 subunit, a major contributor to transient K+ outflow, is targeted by activity-dependent synaptic modulations and its functions are considerably mediated with the activities of synaptic glutamate receptors in CA1 neurons of rat hippocampi [16,23,24]. These previous papers indicate that trafficking properties of Kv4.2 channels are rapidly driven by cellular signalings activated by synaptic activities. Because the rapid internalization of Kv4.2 channels by synaptic activities or plasticities resulted in the reduction of transient currents, the blockade of synaptic activities in CA1 neurons during recording was necessary for testing effects of HpTX2 on K+ outward currents. For this, we used a nucleated-patch technique for isolating soma membrane of a CA1 neuron (Fig. 1A). This methodological approach allowed us to test the intracellular or extracellular effects of HpTX2 without influences of synaptic activities. In all experiments performed in this study, nucleated-patches were completely pulled within 2 min after making whole-cells, and neurons were rejected in cases of over 2 min. In the previous paper, the extracellular application of 100~200 nM HpTX2 blocked only IA currents through Kv4 subunits in rat ventricular myocytes [18]. Therefore, we first tested 200 nM HpTX2, which was added to external ACSF, to determine whether it could block transient currents in CA1 neurons. Total K+ outward currents were recorded at +60 mV holding potential in a voltage-clamp mode, and sustained and transient currents were isolated by using a pre-pulse protocol (Fig. 1A). Consistent with previous reports, the prominent effect of HpTX2 was observed only on transient currents [18,19]. CA1 neurons showed a small but significant reduction of transient current amplitude in the presence of 200 nM HpTX2, compared with neurons tested in normal recording solution (Fig. 1B and C, transient, control=1.57±0.45, n=5; HpTX2=1.32±0.71 nA, n=6; p<0.05). However, sustained currents were not affected by extracellular HpTX2 (Fig. 1B and C, sustained, control=0.19±0.01; HpTX2=0.18±0.01 nA). These results indicate that existence of HpTX2 in extracellular spaces can modulate the gating properties of Kv4.2 subunit in CA1 hippocampal neurons. Extracellular applications in 0.2 to 10 µM of HpTX2 (Supplementary Fig. 1) were also tested for confirming the concentration-dependent effects on CA1 neurons. All concentration of HpTX2 significantly reduced transient currents of CA1 neurons, but no differences of reduction rates according to each concentration were observed. This indicates a restricted modulation of gating kinetics of IA channels by HpTX2 in CA1 neurons. Also, we confirmed that long exposure to HpTX2 (2 µM, 24 hours) exhibited the similar rate of currents reduction, compared with short exposure effects. All concentrations of extracellular HpTX2 showed no effects on sustained currents (Supplemental Fig. 1).
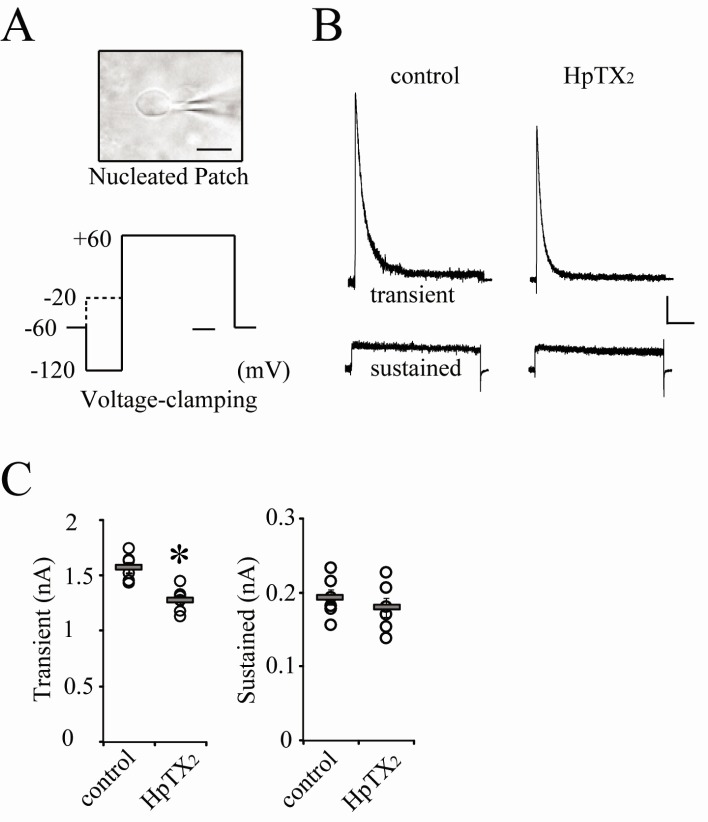
Fig. 1
Extracellular HpTX2 reduces transient K+ outward currents in CA1 neurons. (A) The microscopic IR view of a nucleated-patch and a protocol of voltage-clamping to induce K+ outward currents. The scale bar inserted in the picture indicates 10 µm. Data acquired from experiments in which nucleated-patches were pulled within 2 min after making whole-cells, were used in this study. A pre-pulse (dotted line) to activate only sustained currents was applied for isolating transient currents from total outward currents. In all experiments, peaks of transient and sustained currents were generated at +60 mV holding potential. The time scale bar for voltage-clamping is 100 ms. (B) Example traces of transient and sustained currents recorded at +60 mV clamping in CA1 neurons. Currents were recorded at 10 min after pulling a nucleated-patch with (HpTX2) or without (control) HpTX2 in external solution. Scale bars indicate 0.2 nA and 100 ms. (C) All individual (circles) and averaged (square bars) amplitudes of transient (left panel) and sustained (right panel) currents are plotted for each group (control and HpTX2). Error bars represent standard error of mean (SEM). *: p<0.05, compared with control.
![]()
It is possible that blocking or modulating effects of toxins on gating properties of channels are variable regarding binding affinities between toxins and channels. In Fig. 2, the next experiments were performed in order to determine if intracellular application of HpTX2 also has effects to block only transient currents in neurons. Using internal pipette solution including either 100 or 200 nM HpTX2, nucleated-patches were pulled within 2 min after making a whole-cell. Unexpectedly, both 100 and 200 nM of HpTX2 were effective on sustained but not transient currents. Approximately 30% of the peak amplitude of sustained currents was reduced by intracellular HpTX2 five minutes after making a whole cell (Fig. 2, 200 nM HpTX2; sustained, 0 min=0.19±0.08, 5 min=0.14±0.04, p<0.01; transient, 0 min=1.59±0.11, 5 min=1.52±0.12 nA; n=11). The reduction of sustained currents by HpTX2 seemed not to be dose-dependent, showing a similar decrement between 100 (n=5) and 200 nM treatments (Fig. 2B). Altering intracellular components is critical for determining channel permeability and neuronal responses, and cytosolic space of soma is significantly restricted under a nucleated-patch condition. Therefore, addition of HpTX2 to an internal solution might possibly affect channel conductances without pharmacological actions. For clarifying this issue, we analyzed changes of current density, which was measured by whole-cell patch capacitances (Fig. 2C). All neurons recorded with intracellular HpTX2 (n=16) did not show significant changes of cell capacitances between 0 min (before recording, 19.8±3.2 pF) and 15 min (after recording, 21.1±2.2 pF) after pulling nucleated-patches, so it is clear that the densities of transient and sustained currents reflected the changes of peak amplitudes (Fig. 2C, transient: 0 min=81.69±3.79, 15 min=74.81±4.02, p=0.248; sustained: 0 min=9.62±0.35, 15 min=6.54±0.29 pA/pF, p<0.01). In addition, we tested other agents known as blockers of K+ channels to determine if they also have intracellular effects on either transient or sustained currents. 4-AP is widely used for suppressing K+ outward currents in dendrites and soma, and was previously confirmed to be effective on transient currents when it was added to internal pipette solution [13,16]. In this study, 3 mM of 4-AP was added to internal solution, and nucleated-patches were then made (Fig. 3A). 4-AP application resulted in a rapid and significant reduction of the peak amplitude in transient currents but not in sustained currents (Fig. 3A, normalized amplitude at 10 min compared with 0 min; transient=0.35±0.04, p<0.05; sustained=0.86±0.02, n=5). We also tested Cs-gluconate internal solution (Fig. 3B). In this experiment, both transient and sustained currents showed a gradual and significant reduction 10 min after pulling nucleated-patches (Fig. 3B, Normalized amplitude at 10 min; transient=0.67±0.04, p<0.05; sustained=0.71±0.03, p<0.05, n=5). Therefore, the results shown in Fig. 2 and 3 suggest that blocking effects of agents tested in this study may possibly be variable according to the location of channel effective sites [18,19]. In Fig. 4, the reduction rates of transient and sustained currents by intracellular drug treatments, which were calculated by the ratio between 0 min and 15 min peak after pulling nucleated-patches, were compared. The reduction rate of HpTX2 in sustained currents was significantly larger than that in transient currents, while 4-AP reduced only transient currents (Fig. 4, HpTX2, transient:sustained=6.43±2.55: 27.68±3.22%; 4-AP, transient:sustained=64.15±3.51:18.02±6.08%). The reduction rates of Cs-gluconate internal sol. were statistically significant in both transient and sustained currents, (Fig. 4, Cs-G. internal, transient-sustained=33.03±3.67:33.29±3.59%). These results suggest the possibility that K+ outward flows in CA1 neurons can be selectively blocked by chemicals or toxins according to their application methods.
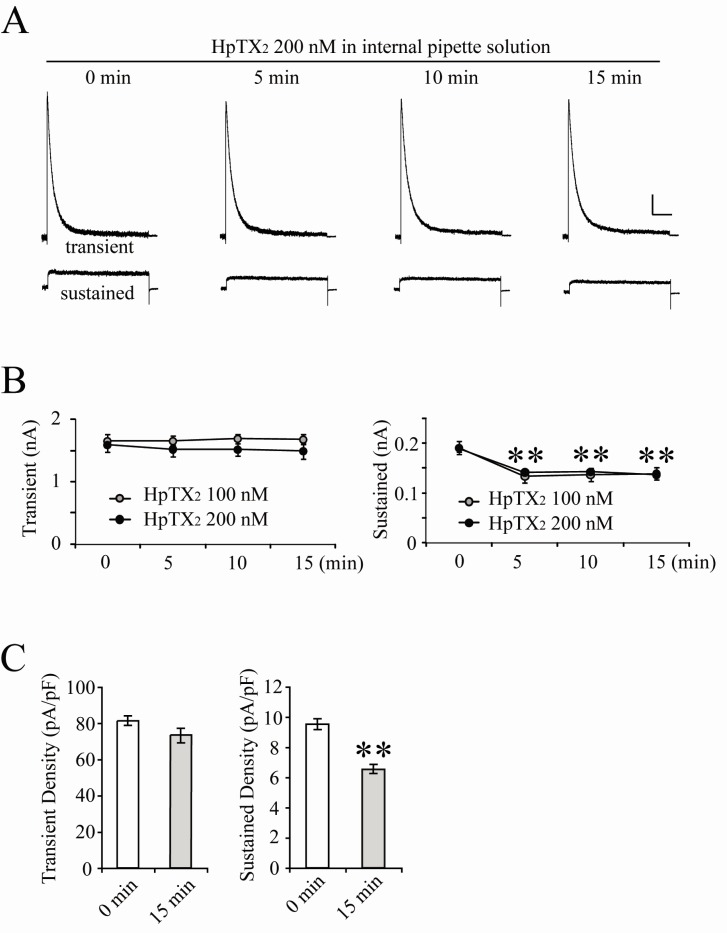
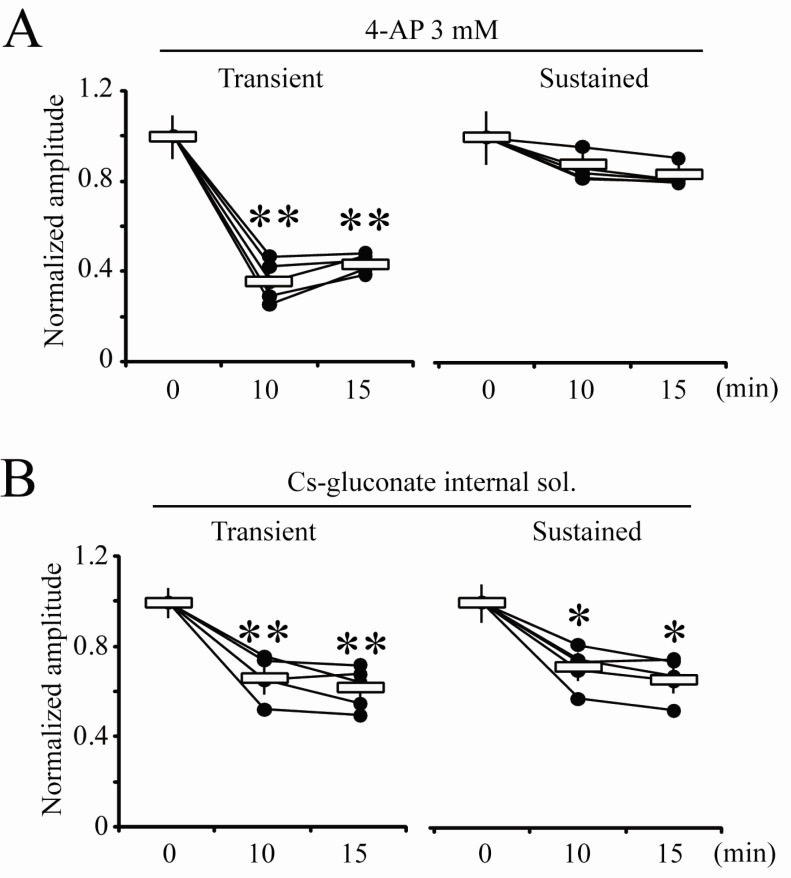
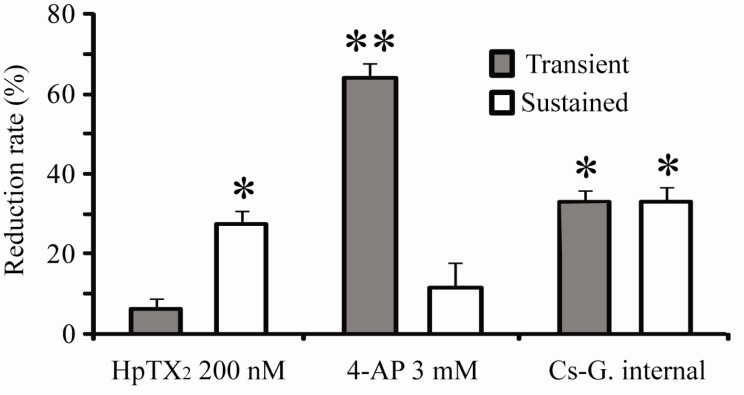
Fig. 2
Cytosolic HpTX2 reduces sustained but not transient K+ outward currents in CA1 neurons. (A) Example traces of both currents recorded immediately (0), 5, 10 and 15 min after pulling nucleated-patches. HpTX2 (200 nM) was added to internal pipette solution and normal external solution was perfused during recording. Scale bars indicate 0.2 nA and 100 ms. (B) Averaged peak amplitudes of transient (left panel) and sustained currents (right panel) recorded with intracellular 100 (gray circles) or 200 nM (dark circles) HpTX2. (C) Averaged current densities of transient and sustained currents measured at 0 min and 15 min after making nucleated-patches. The changes of current densities were acquired from data shown in (B) (100 nM: n=5, 200 nM: n=11). To calculate current density, the whole cell capacitance of 0 min was checked before recording currents, and that of 15 min was checked again right after completing the recording. Error bars represent SEM. **: p<0.01, compared with 0 min.
![]()
Fig. 3
Blocking effects of cytosolic 4-AP and Cs-gluconate internal solution on K+ outward currents. (A) Normalized individual and averaged values of peak amplitudes of transient and sustained currents recorded with 4-AP (3 mM) in internal pipette solution. (B) Normalized individual and averaged values of peak amplitude of transient and sustained currents recorded with Cs-gluconate internal solution. Error bars represent SEM. *: p<0.05 or **: p<0.01, compared with 0 min.
![]()
Fig. 4
Comparison of blocking effects on transient (gray bar) and sustained (open bar) currents induced by cytosolic drugs in hippocampal CA1 neurons. The reduction rate was calculated by the ratio of decreased amplitudes at 15 min after pulling nucleated-patches, compared with "0 min" peak amplitudes. Error bars represent SEM. *: p<0.05 or **: p<0.01, compared with "0 min" values shown in Fig. 2 and 3.
![]()
DISCUSSION
The opening properties and current flows through K+ channels are critical factors in regulation of neuronal excitabilities. Therefore, studying agents to block specific subunits of K+ channels is valuable for understanding neuronal functions. Major findings in this study are that HpTX2 is not a specific agent to block or modulate transient currents, and that the intracellular application of HpTX2 can reduce sustained K+ outward currents in CA1 hippocampal neurons. Although these results are not consistent with previous reports demonstrating HpTX2 effects to be specific on Kv4 families, it should be considered that sustained and transient currents were sensitive to intracellular and extracellular HpTX2, respectively. Because testing HpTXs effects has previously and dominantly been performed in Xenopus oocytes or cardiac myocytes, the effective reduction in both sustained and transient currents in CA1 neurons of hippocampal slices indicates that HpTX2 is also valuable for studying the physiological functions of K+ channels working in neuronal excitabilities of mammalian CNS.
Transient currents through Kv4.2 channels in CA1 neurons play crucial roles in modulation of AP back-propagation, dendritic integration of excitatory postsynaptic potentials, synaptic composition of glutamate receptors and intrinsic excitabilities [13,16,24,25]. In particular, synaptic activities of hippocampal neurons can directly regulate Kv4.2 trafficking in dendrites and soma [23]. Therefore, most studies investigating modulatory functions of K+ channels in neuronal excitability, primarily focus on either Kv4.2 or Kv1.4 subunit. However, pharmacological approaches to block Kv4 and Kv1 families have difficulties in the absence of specific chemical agents, even though some chemicals were useful for blocking the various K+ outward currents in cardiac and neuronal cells [13,26,27]. In the last decade, pharmacological studies of some toxins from insects and snakes have suggested a possibility for the manipulation of specific Kv subunits, and provided credible results that HpTXs and hanatoxins are valuable to determine neuronal functions of Kv channels [12,18-20]. Of them, HpTX2 is known as a gating regulator specific to Kv4 subunits via binding to the S3-S4 linker region of Kv4.2 [19]. However, because of insufficient evidence in mammalian CNS, it is not suitable for determining pharmacological effects of HpTX2 on native channels in neurons. In addition, rapid changes in trafficking and gating properties of Kv4 subunits are induced by synaptic activities [16,23]. In this study, extracellular application of 200 nM HpTX2 resulted in a small but significant reduction of transient but not sustained K+ outward currents in nucleated-patched CA1 neurons. This indicates that HpTX2 can act as a gating modulator of Kv4.2 subunit natively expressed in CA1 neurons, consistent with effects on channels expressed experimentally in Xenopus oocytes [18,19]. However, intracellular HpTX2 significantly reduced sustained currents, showing no changes in transient currents. Compared with intracellular 4AP that reduced only transient currents, it is possible that the blocking effect of cytosolic HpTX2 on sustained currents may be due to pharmacological interactions with Kv2.1 subunit showing homologue structures with other Kv families [22]. Because hydrophilic or hydrophobic property of chemicals is correlated with their membrane permeability, different actions between intracellular and extracellular HpTX2 can be explained by a hypothesis that hydrophilic HpTX2 and the location of effective sites in channels may determine the toxin effects on Kv channels [19].
Kv2.1 channels, as well as Kv4 families, are major determinants of neuronal excitabilities, so they can regulate neuronal responses based on synaptic inputs and dendritic integrations. In addition, several reports have provided evidence that sustained K+ currents can continuously compensate for activity-dependent changes of transient currents and membrane excitabilities via regulating intracellular K+ concentrations [28-31]. Therefore, distinctive actions of HpTX2 inside and outside of neurons are very effective for the selective reduction of specific K+ outward currents, compared with effects of Cs-gluconate internal solution acting in both transient and sustained currents. In the present study, we did not confirm the interaction mechanism between intracellular HpTX2 and channels contributing to sustained currents. The precise actions of cytosolic HpTX2 to correlate with Kv subunits should be addressed in a future study.
 XML Download
XML Download