

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Most circulating blood cells are derived from hematopoietic stem cells (HSCs), which have various functions including self-renewal and differentiation into specialized cells. HSCs mature into specific organ cell type and are periodically replenished by new HSCs in their niche during the life span of an animal. Moreover, HSCs are known to have 2 critical properties, multipotency and self-renewal, that can lead to their differentiation into multiple lineages and preservation of HSC fate upon cellular division, respectively [1]. The HSC life span is regulated by various internal and external signals mediated by stimuli such as transcription factors, growth factors, and cell cycle regulators [2,3]. Many researchers have tried to identify various methods, such as the use of a combination of cytokines or infection with specific genes, to improve HSC survival, proliferation, and self-renewal [4,5]. Gene members of the Hox A, B, and C clusters are reported to be expressed in hematopoietic cells [6]. Thus, the Hox family has gained attention because of its involvement in the functional regulation of HSCs. However, the pro-B cell line Ba/F3, an important model system in cancer research that is interleukin-3 (IL-3) dependent for its proliferation and survival [7,8], remains unexamined.
Hox is considered a key regulator that controls the fate of hematopoietic stem and progenitor cells [9,10]. The Hox family harbors a highly conserved DNA-binding domain (homeodomain) and induces transcriptional regulation via interaction with a variety of transcriptional co-factors (for example, Pbx and Meis) to achieve specific transcriptional programming throughout hematopoietic development [10-13]. Among homeobox transcription factors, HoxB4 is known to be involved in the regulation of normal and leukemic hematopoiesis at various stages. HoxB4 was reported as one of the factors that can independently facilitate the expansion of HSCs when expressed ectopically [13]. Moreover, retroviral delivery of HoxB4 mediates the expansion of murine long-term reconstituting HSCs [13-15]. Milsom et al. have demonstrated that HoxB4 suppresses multiple components of the tumor necrosis factor-α (TNF-α) signaling pathway by direct transcriptional regulation of TNF-α effectors [16].
Recently, it has been reported that HoxB4 influences the expression of genes involved in intrinsic pathways such as cell cycle regulation, differentiation, and apoptosis; in particular, the ectopic expression of HoxB4 modulates Bre transcripts, which are involved in HSC self-renewal and in the inhibition of the Fas-associated cell death signal [17]. However, the effects of HoxB4 on biological events in other cells have not been examined yet. Therefore, in this study, we investigated whether HoxB4 gene overexpression affects the survival of the murine pro-B cell line Ba/F3 in various death-inducing environments using apoptosis and the cell cycle assay. We demonstrate in the present study that the constitutive expression of the HoxB4 gene may be directly or indirectly related to pro-B cell survival, which may result in maintenance of the survival state or postponement of apoptosis and necrosis under extreme conditions.
Go to : 
METHODS
Cell culture
Cells from the murine pro-B cell line Ba/F3 were cultured in Roswell Park Memorial Institute 1640 (RPMI 1640; Gibco BRL, Rockville, MD, USA) medium supplemented with 10% fetal bovine serum (FBS; Gibco BRL), penicillin/streptomycin (25 µg/ml), and IL-3 (5 ng/ml; Peprotech, Rocky Hill, NJ, USA). 293GPG cells were maintained in Dulbeco's modified eagle medium (Gibco BRL) with 10% FBS, penicillin/streptomycin (25 µg/ml), G418 (300 µg/ml), puromycin (2 µg/ml), and tetracycline (1 µg/ml).
Generation of retroviral vector
The pGCDNsam-IRES-GFP (MSCV) retroviral vector used in this study was kindly provided by Dr. Onodera from University of Tsukuba (Tsukuba, Ibaraki, Japan). The MSCV-GFP and MSCV-HoxB4 retroviral vectors were based on the MSCV backbone. The pGCDNsam-HoxB4 plasmid contains the murine HoxB4 cDNA followed by the encephalomyocarditis virus internal ribosome entry site (IRES) and the gene encoding enhanced green fluorescent protein (GFP). HoxB4 cDNA from murine bone marrow was obtained using polymerase chain reaction (PCR) with specific primers (5'-AGGCGGATCCTCCCA GAAATTAATGGC TATGAG and 3'-TAATCTCGAGCCCCTTCGGTGCTCGCT GTT C), and inserted into the BamH1 and Xho1 sites of MSCV. A high-titer-producing stable cell line was generated via transfection with the MSCV-HoxB4 and MSCV-GFP control vectors into 293GPG cells using lipofectamine (Invitrogen, Carlsbad, CA, USA) in accordance with the manufacturer's instructions, and GFP-positive cells were sorted via fluorescence-activated cell sorting (FACS) (FACS Vantage SE; BD Bioscience, Mansfield, MA, USA). Following this, we added retroviral supernatants acquired from 293GPG-GFP and 293GPG-HoxB4 cells to cultures of Ba/F3 cells. After 2 days, GFP-positive cells were obtained using FACS (BD Bioscience).
Reverse transcription polymerase chain reaction (RT-PCR) analysis
Total mRNA was isolated from Ba/F3-GFP and Ba/F3-HoxB4 cells using the TRIzol reagent (Gibco BRL). These RNAs were used as templates for RT-PCR using the HoxB4 primers mentioned above. PCR was performed at 94℃ for 3 min, 55℃ for 30 sec, and 72℃ for 1 min for a total of 30 cycles. β-actin was utilized as an internal control.
Cell proliferation and cell cycle assay
To determine the cell proliferation capacity of HoxB4, Ba/F3 cells infected with HoxB4 and GFP control vector were washed twice in 1× PBS. Cells were seeded in 96-well plates at a density of 2×104 cells per well in a final volume of 100 µl RPMI without IL-3, and cultured for 3 days after seeding. Cells were treated with 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) reagent and the absorbance at 595 nm was measured by enzyme-linked immunosorbent assay (ELISA).
To determine the effects of HoxB4 on the cell cycle, 5×105 Ba/F3-GFP or Ba/F3-HoxB4 cells were incubated in media with or without 100 nM doxorubicin (DOX) for 24 h. After harvesting and fixing in 70% ethanol, the cells were treated with 50 µg/ml RNase in 1× PBS for 5 min at room temperature. Next, 50 µg/ml propidium iodide (PI) was added for 10 min at 37℃ followed by FACS analysis.
Apoptosis assay
To assess the effects of DOX on cell death, cells were incubated with an agonistic anti-Fas antibody (clone Jo2, BD Bioscience) with or without 100 nM DOX for 24 h. The annexin V assay (BD Bioscience) was employed to detect phosphatidylserine (PS) exposure on the outer-leaflet of the plasma membrane. Early and late apoptosis were measured by double staining with PI and annexin V-phycoerythrin (PE). 5×105 cells were seeded in 24-well plates in media with or without 100 nM DOX. After 24 h, the cells were washed with phosphate-buffered saline (PBS) and resuspended in binding buffer (100 mM Hepes/NaOH, pH 7.4; 140 mM NaCl; 2.5 mM CaCl2). Annexin V (1 µg/ml) and PI (1 ng/ml) were added, and the cells were then incubated in the dark at room temperature for 15 min. The cells were washed with binding buffer and analyzed by flow cytometry.
Flow cytometry analysis
Ba/F3-GFP and Ba/F3-HoxB4 cells were plated in 6-well plates (5×106 cells in each well) and incubated with or without 100 nM DOX for 24 h. Cells were then harvested and washed 2 times with PBS. Cells were stained with biotinylated Fas antibody (clone DX2; PharMingen, Torrey Pines, CA, USA) or an isotype-matched control rat anti-mouse IgG1 antibody (PharMingen), and then labeled with streptavidin-conjugated allophycocyanin (APC). The cells were also incubated with PI to exclude dead cells, and analyzed by flow cytometry. To investigate alterations in morphology, Ba/F3-GFP and Ba/F3-HoxB4 cells were examined by flow cytometry, variable cells were detected using 488-nm light from an argon laser, and the forward scatter (FSC) and side scatter (SSC) were determined.
Data analysis
The statistical significance of the differences between experimental groups was evaluated via two-tailed Student's t test. Differences of p<0.05 were considered to be statistically significant.
Go to :
RESULTS
HoxB4 overexpression inhibits the expansion of cell size and granularity in Ba/F3 cells
In order to understand the effect of HoxB4 on cell morphology, we first overexpressed HoxB4 in Ba/F3 cells. As shown in Fig. 1A, HoxB4 was cloned into a retroviral vector (MSCV viral backbone) with the long terminal repeat (LTR) and GFP regions (HoxB4-GFP virus). The analogous vector harboring GFP alone (GFP virus) was utilized as a control. After checking that GFP and HoxB4 were stably expressed in Ba/F3 cell lines by sorting GFP-positive cells using flow cytometry, ectopic HoxB4 overexpression was confirmed via RT-PCR analysis using HoxB4 specific primers (Fig. 1B). Next, the effect of HoxB4 overexpression on the morphology of cells was tested using FACS analysis and microscopic observation. In the FACS analysis, HoxB4 overexpression decreased cell granularity (side scatter; SSC) and size (forward scatter; FSC) in Ba/F3 cells compared with the control (Fig. 1C). Similarly to this result, in the microscopic observation, HoxB4-overexpressing cells showed predominant morphological change compared with the control (Fig. 1D).
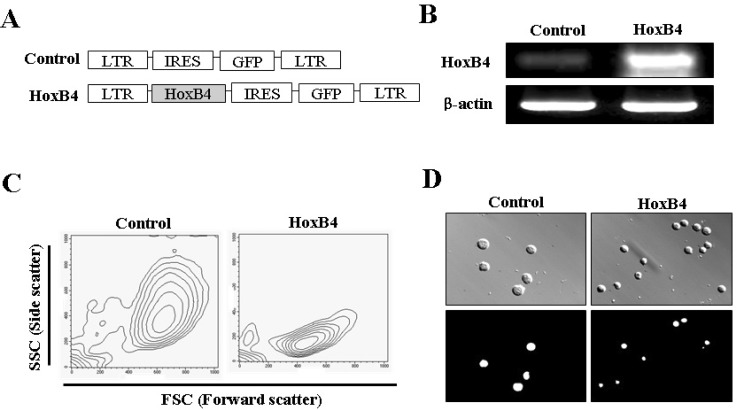 | Fig. 1Retroviral vectors and phenotype induced by HoxB4 in Ba/F3 cells. (A) Constructs of MSCV-GFP and MSCV-HoxB4 retroviral vectors. Schematic representation of the MSCV-HoxB4-IRES-GFP construct and the MSCV-IRES-GFP control vector are shown. (B) Confirmation of HoxB4 expression in Ba/F3 cells. Using cDNA from both infected cells, we examined the effect of HoxB4-expressing Ba/F3 cells using reverse transcription-polymerase chain reaction (RT-PCR). (C) Phenotype of Ba/F3-HoxB4 cells. The morphology of the HoxB4-infected cells was demonstrated via FSC (forward scatter) and SSC (side scatter) in fluorescence-activated cell sorting (FACS) analysis. (D) Photomicrograph of Ba/F3 cells with HOXB4 overexpression. Upper panels, cell images observed under condition of phase contrast illumination; Lower panel, GFP fluorescence images in cells corresponding to phase contrast images. Representative pictures are shown from at least three independent experiments. Magnification ×400.
|
HoxB4 promotes cell proliferation and suppresses apoptosis in the absence of survival signals
Based on the induction of different phenotypes in Ba/F3 cells by HoxB4 overexpression, we tested various biological events in Ba/F3-HoxB4 cells. Analysis of the cell proliferation response using the MTT method revealed a significant increase in proliferation in Ba/F3-HoxB4 cells compared with Ba/F3-GFP cells in a time-dependent manner (Fig. 2A). Cell cycle analysis showed that the proportion of Ba/F3-GFP cells in the G0/G1 and G2/M phases were 51.3% and 19.6%, respectively, whereas the corresponding proportions in Ba/F3-HoxB4 cells were 46.6% and 20.1%, respectively, implying that HoxB4 reduced the number of cells in the G1/G0 phase. The number of Ba/F3-HoxB4 cells in the S-phase tended to be slightly higher than the number of Ba/F3-GFP cells in the same phase (Fig. 2B). The number of Ba/F3-HoxB4 cells in the sub-G1 phase was 15.4-fold lower than the corresponding number of Ba/F3-GFP cells. The survival of Ba/F3 cells is known to be dependent on the IL-3 signal, and IL-3 deprivation leads to cell death via the activation of pro-apoptotic members of apoptosis [8]. We therefore assessed apoptosis levels in IL-3-deficient Ba/F3-HoxB4 cells to investigate cellular events induced by the removal of the survival signal. Both Ba/F3-GFP and Ba/F3-HoxB4 cells were cultured for 1 day in conditions of starvation with the removal of IL-3, and then the extent of apoptotic cell death was measured using annexin V and PI analysis. We observed that HoxB4 overexpression decreased the level of apoptotic and necrotic cells from 64.6% to 54.1% (Fig. 2C). Interestingly, the level of necrotic and late apoptotic cells declined from 51.2% to 30.8%. These results imply that HoxB4 overexpression in pro-B cells inhibits cell growth and cell death, resulting in an increase in the population of Ba/F3 cells.
 | Fig. 2Biological activities of HoxB4 in Ba/F3 cells. (A) Proliferation rate of Ba/F3-HoxB4 cells. 2×103 Ba/F3-HoxB4 cells were cultured with 5 ng/ml IL-3 (survival cytokine) in 96-well plates for 3 days and cell proliferation activity was assessed via MTT assay. Data are expressed as mean±standard deviation (SD). *p<0.05 in comparison with control cells. (B) Alteration of the cell cycle by HoxB4. Ba/F3-GFP and Ba/F3-HOXB4 cells were stained with PI and detected by fluorescence-activated cell sorting (FACS) after incubation in normal culture conditions. Data are expressed as mean±SD. (C) Inhibitory effect of apoptotic cell death by HoxB4. Ba/F3-GFP and Ba/F3-HoxB4 cells were incubated without IL-3 (-) for 24 h, and then the apoptotic cell death of Ba/F3-HoxB4 cells was measured using the Annexin V and PI assay with FACS. The data express the rate of double-negative (live), single-positive (early apoptosis and necrosis), and double-positive (late apoptosis) cells.
|
HoxB4 prevents apoptotic cell death under conditions of cell cycle arrest in G2/M phase
It is reported that DOX acts as an anticancer therapeutic that induces cell death via cell cycle arrest in the G2/M phase, and exerts stronger effects against rapidly growing cells [18]. To examine the effect of HoxB4 on these cell responses in Ba/F3 cells, we analyzed apoptotic cell death and cell cycle arrest in Ba/F3-GFP and Ba/F3-HoxB4 cells treated with DOX. Analysis of cell proliferation and the cell cycle after 24 h of incubation in 100 nM DOX revealed that 76.1% of Ba/F3-HoxB4 and 48.4% of control cells were arrested in the G2/M phase (Fig. 3A). The number of Ba/F3-HoxB4 cells in the sub-G1 phase was lower than that of control cells, suggesting that HoxB4 may be associated with the prevention of apoptotic cell death, although it causes a dramatic cell cycle arrest at G2/M phase.
 | Fig. 3Biological effects of HoxB4 in cell cycle arrest at G2/M phase. (A) HoxB4 in the cell cycle. Ba/F3-HoxB4 cells were incubated with 100 nM DOX for 24 h, and then the DNA content was determined by PI staining with flow cytometry. Representative flow cytometry data and the percentage of cells at each stage of the cell cycle are shown. These data represent the mean±standard deviation (SD) from 3 independent experiments. *p<0.05 in comparison with control cells. (B) Apoptosis assay of Ba/F3 cells infected with HoxB4. Ba/F3-HoxB4 cells were incubated with media containing 100 nM DOX. After staining with Annexin V-PE and PI, we performed flow cytometry. The data represent the proportions of double-negative (live), single-positive (early apoptosis and necrosis), and double-positive (late apoptosis) cells.
|
In addition, to examine whether HoxB4 affects cell death induced by cell cycle arrest, we treated Ba/F3-GFP and Ba/F3-HoxB4 cells with 100 nM DOX for 24 h, and then analyzed apoptotic cell death by staining the cells with annexin V-PE and PI. As shown in Fig. 3B, Ba/F3-HoxB4 cells showed slightly decreased cell death compared to control cells. The survival rates of Ba/F3-HoxB4 and Ba/F3-GFP cells were 79.6% and 77.4%, respectively, and the apparent progression of cell death was slower in Ba/F3-HoxB4 cells than in control cells (Fig. 3B). These results suggest that HoxB4 may be directly involved in cell cycle progression but does not have a significant effect on cell death by cell cycle arrest.
HoxB4 prohibits apoptotic cell death by Fas stimulation
Recently, we reported that DOX arrests the cell cycle at G2/M phase and increases Fas expression on the cell surface [19]. We therefore tested the effect of HoxB4 overexpression on the alteration of Fas expression by DOX treatment. As shown in Fig. 4A, when treated with 100 nM DOX, the intensity of Fas expression in Ba/F3-HoxB4 cells was increased from 82.1 to 435.1 and that in control was increased from 71.7 to 291.6. This result indicates that HoxB4 may enhance the increase in Fas expression induced by DOX treatment.
 | Fig. 4Biological activities of HoxB4 in conditions that stimulate the Fas (CD95) apoptotic signaling pathway. (A) Ba/F3-GFP and Ba/F3-HoxB4 cells were cultured with 100 nM DOX for 24 h, recovered, and stained with anti-Fas (CD95) antibody, and immune fluorescence was quantified by flow cytometric analysis. Numbers in the upper parts of histogram curves indicate the positive intensity values of Fas expression in cells. The graph was obtained from left two panels and represents fold changes between results from Ba/F3-GFP and Ba/F3-HoxB4 cells in the presence of DOX. *p<0.05 in comparison with control cells. (B) Stimulation of the Fas apoptotic signal pathway. Ba/F3-GFP and Ba/F3-HoxB4 cells were treated with 1 µg/ml agonistic antibody (clone Jo2) for 24 h and analyzed by flow cytometry after staining with Annexin V-PE and PI. The data represent the proportions of double-negative (live), single-positive (early apoptosis and necrosis), and double-positive (late apoptosis) cells. (C) Stimulation of Fas induced by DOX. Ba/F3-GFP and Ba/F3-HoxB4 cells were treated with 1 µg/ml agonistic antibody (clone Jo2) and 100 nM DOX for 24 h and analyzed by flow cytometry after staining with Annexin V-PE and PI. The data express the proportions of double-negative (live), single-positive (early apoptosis and necrosis), and double-positive (late apoptosis) cells. (D) Photomicrograph of cells incubated with DOX and Jo2 antibody. Ba/F3-GFP and Ba/F3-HoxB4 cells were seeded in 24-well plates (5×105 cells in each well) and incubated with complete media, 100 nM DOX or 1 µg/ml Jo2 antibody and after 24 h, the morphology change of cells were observed in each well. Magnification ×40.
|
To investigate the effects of HoxB4 on apoptotic cell death by Fas stimulation, we analyzed the levels of annexin V and PI in Ba/F3-HoxB4 cells following treatment with Jo2 (Fas agonistic) antibody and DOX. As shown in Fig. 4B, Ba/F3-HoxB4 cells that were treated with Jo2 antibody showed an increase of about 29.2% in double-negative (surviving) cells and a decrease of almost 20% in apoptotic cell death compared with control cells. On the other hand, annexin-V- and PI-positive Ba/F3-GFP cells also significant induced cell death (21.9%), in contrast to PI-positive Ba/F3-HoxB4 cells (5.5%). The proportion of early apoptotic cells was 26.9% in Ba/F3-GFP cells and 15.1% in Ba/F3-HoxB4 cells. The proportion of late apoptotic cells was 9.7% in Ba/F3-GFP cells and 1.5% in Ba/F3-HoxB4 cells. These results suggest that HoxB4 may be associated with cell survival via the Fas-mediated apoptotic signaling pathway. Moreover, Ba/F3-HoxB4 cells, stimulated by DOX-induced Fas, showed a greater reduction in apoptotic cell death than did Ba/F3-GFP cells. The number of early apoptotic Ba/F3-HoxB4 cells was decreased by 2.7%, whereas that of late apoptotic cells was reduced by 11.2% and that of necrotic cells was diminished by 7.3% (Fig. 4C). Furthermore, in the direct cell counting analysis, HoxB4-overexpressing cells showed to exert inhibitory effect on cell death induced by treatment with Jo2 antibody and DOX (Fig. 4D). These results indicate that HoxB4 can prevent apoptotic cell death by Fas stimulation, although it enhances the increase of Fas expression on the cell surface.
Go to :
DISCUSSION
It is reported that HoxB4 plays key roles in HSCs and in several hematopoietic progenitor cells including granulocyte macrophages and lymphoid progenitors [15,20]. It is a novel growth stimulatory regulator of primitive hematopoietic cells [13] and can rapidly expand in vitro cell populations, including NOD/SCID repopulating cells, LTC-ICs, and committed clonogenic progenitors [15]. Although many studies on the function of HoxB4 have been reported, the biological effects of HoxB4 in Ba/F3 cells have not been examined yet. Thus, in the present study, we investigated the survival effects of HoxB4 in various conditions of cell death, such as cell cycle arrest or stimulation of Fas protein expression in pro-B cells following the overexpression of HoxB4 genes and the removal of survival factors.
In the present study, we were able to draw three major conclusions from our results. First, HoxB4 not only inhibits cell growth and cell death but also increases the rate of cell proliferation. Transfection with HoxB4 dramatically decreased cell size and granularity compared to control cells. In addition, transfection with HoxB4 significantly increased proliferation and the S phase of the cell cycle, and attenuated natural apoptotic cell death under normal culture conditions. Therefore, we suggest that HoxB4 may play important roles in cell growth, proliferation, and survival. Second, HoxB4 strengthens survival ability, as well as rapid cell proliferation. In the present study, we examined whether HoxB4 transfection into Ba/F3 (an IL-3 cytokine-dependent B cell line) cells affects apoptosis by removing IL-3 (survival growth factor) from the culture media or by adding DOX, which arrests the cell cycle in the G2/M phase. We found that Ba/F3-HoxB4 cells showed progressive attenuation of cell death from early to late apoptosis. These results indicate that HoxB4 extends apoptotic progression in pro-B cells via the interception of the cell survival signaling pathway and the induction of cell cycle arrest in the G2/M phase. Finally, we investigated whether HoxB4 expression overcomes the inhibitory effects of the apoptotic Fas proteins in the presence or absence of DOX. In our previous study, we showed that DOX induced significant overexpression of the Fas protein, which is known as a proapoptotic protein on the cell surface of Ba/F3, and EL4 cells, and in T cell lines [19]. Moreover, Fas proteins expressed upon DOX treatment have apoptotic activities, which were validated by Annexin V-PE and PI assays. In the present study, Fas expression in Ba/F3-HoxB4 cells treated with 100 nM DOX was significantly higher than that of Ba/F3-GFP cells, indicating that Ba/F3-HoxB4 cells were more affected than Ba/F3-GFP cells by a Fas apoptotic signal. However, the apoptotic effect of Fas proteins, which was stimulated by treatment with DOX and Jo2 (Fas agonistic) antibodies, was attenuated upon HoxB4 expression. Therefore, our data suggest that HoxB4 has the important function of protecting against the apoptotic signal by shutting off the survival signal, arresting the cell cycle in G2/M phase, and attenuating Fas signal stimulation.
It is reported that HoxB4 influences the expression of genes involved in pivotal intrinsic regulation pathways of cellular responses such as cell cycle, differentiation, and apoptosis [14]. Moreover, induction of HoxB4 decreases the expression of p21, p27, cyclin G2, and cyclin M2, and increases the expression of cyclin D1 [17]. We can therefore conjecture that decreased p21 gene expression suppresses cell death, because p21 is specially cleaved by CASP3-like caspase, which leads to dramatic activation of CDK2, and is important for the induction of apoptosis following caspase activation [21-23]. In addition, p27 and cyclin G1 are known to inhibit cell proliferation and modulate the cell cycle and cellular division processes [24,25]. It was demonstrated that HoxB4 can induce expression of cyclin D1 proteins that regulate CDK kinases and interact with CDK4 or CDK6, whose activity is required for the cell cycle G1/S transition [26]. These reports imply that increased HoxB4 may accelerate cell proliferation and cell cycle progression by diminishing the expression of p21, p27, and cyclin G2, and by increasing cyclin D1 expression. It is also assumed that HoxB4 up-regulates various transcription factors such as c-myc, Jun-B, and Fra-1 [27], and that this response may promote cell proliferation and replication. HoxB4 is reported to lead to reduced transcriptional levels of the GADD45a gene under stressful growth arrest conditions or following treatment with DNA-damaging agents [28,29]. Further, Bre genes can down-regulate apoptosis by binding the intracellular domain of TNF receptor 1 (TNFR1) and Fas, which is regulated by HoxB4, and inhibit the activation of the mitochondrial apoptotic pathway [30]. Therefore, we suggest that HoxB4 may prevent the progression of apoptotic cell death by up-regulating Bre transcription and down-regulating GADD45a, when these cells are cultured under conditions of cell death such as the removal of IL-3 survival factor, cell cycle arrest by DOX, or the Fas stimulation signal, although these events were not directly investigated in the present study.
In conclusion, the present study demonstrated that the HoxB4 gene in pro-B cells may inhibit the apoptosis and necrosis induced in extremes of cell death, in addition to promoting cell proliferation and limiting cell size. Thus, we suggest that HoxB4 may be a key gene responsible for cell proliferation and survival in Ba/F3 cells. However, further study is required to elucidate the detailed mechanism of the involvement of HoxB4 in cell proliferation and survival in Ba/F3 cells.
Go to :
 XML Download
XML Download