

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Acute ischemic stroke is one of the leading causes of adult disability and death in the world, and it affects up to 0.2% of the population every year [1]. Neuronal cell injury, including apoptosis, is the major event in the acute and sub-acute phases of cerebral ischemia [2,3]. Although the exact mechanism of the neurological destruction of the brain caused by cerebral ischemia has not yet been determined, several studies have suggested that oxidative stress, inflammatory cytokines and excitotoxicity may play a role in etiology of this disease [4,5].
Ginseng (Panax ginseng C.A. meyer, Araliaceae) is a traditional medicinal herb, and it is considered to be an adaptogen that improves physiological function [6,7]. In Asia there are a variety of commercial ginseng products, including white, red and black ginseng. White ginseng is produced as a result of dehydration by the sun, while red ginseng is produced by steaming at 95~100℃ for 2~3 h. Black ginseng is produced from red ginseng by 9 cycles of steam treatment, at which point it becomes black in color [8,9]. The pharmacological and biological activities of steam-processed ginseng are greater than that of non-steamed ginseng. During the steaming process, the ratio of major components, including ginsenosides, saponin, phenolics and proteins, is altered as newly produced active components [10,11]. It has been suggested that the steamed ginseng has a greater spectrum of pharmacological effects than that of white ginseng [12-14], including anti-inflammatory, anti-carcinogenesis, anti-stress and antioxidant effects in humans and animals [8,14-16]. Song et al. and Sun et al., reported that the data of HPLC analysis of BG [9,17]. However, few papers on BG products efficacy have been published.
The present study was undertaken to evaluate the neuroprotective effect of BG on the ischemia-induced learning and memory deficits in rats and to elucidate the cholinergic and neural nitric oxide synthesis mechanism underlying these protective effects in rats.
Go to : 
METHODS
Animals
Male Sprague-Dawley rats weighting 250~280 g each were purchased from Samtaco Animal Corp. (Kyungki-do, Korea). The animals were allowed to acclimatize themselves for at least 7 days prior to the experimentation. The animals were housed in individual cages under light-controlled conditions (12/12-hr light/dark cycle) and at a 23℃ room temperature. Food and water were available ad libitum. All the experiments were approved by the Kyung Hee University Institutional animal care and use committee. The rats were allowed at least 1week to adapt to their environment before the experiments.
Preparation of the black ginseng (BG)
All the 6-year Korean white and red ginseng roots (P. ginseng C.A. Meyer) were purchased from the largest Korean ginseng market of Geumsan country (Chungcheong Province, South Korea). BG was manufactured by repeatedly steaming WG at 98℃ for 3 h in a pottery apparatus and drying it at 60℃ for 15 h. The dried BG samples (40 g) were immersed in a 200 ml of dH2O and added 2 L of 80% ethanol and then mixed 500 ml of dH2O. For the extraction, diethyl ether was used three times.
Middle cerebral artery occlusion model
Focal cerebral ischemia was induced using the intraluminal filament technique. Anesthesia was administered with 3% isoflurane in 30% O2/70% N2O, and it was maintained throughout the operation with 0.5~0.6% isoflurane delivered via a nose mask. The right common carotid artery was exposed through a midline cervical incision. A heparinized intraluminal filament (φ0.28 mm, rounded tip) was introduced via the external carotid artery. The rectal temperature was monitored and maintained at 37℃ using a heating pad (Harvard Homeothermic Blanket Control Unit, 50-7061). After 120 min of occlusion, the filament was gently pulled out and the external carotid artery was permanently closed by cauterization. In the sham-operated rats, the right common carotid artery was exposed and the external carotid artery was opened without introducing the filament into the internal carotid artery. After the operation, the animals were allowed to wake up in the incubator (30℃) and they were then moved to their home cages. The rats were divided into four groups. The experimental group was treated with BG100 [100 mgkg-1, p.o., BG100+ISCH group (n=6)] and BG400 [400 mgkg-1, p.o., BG400+ISCH group (n=6)] for 2 weeks after the induction of ischemia. The control group was treated with saline [0.1 mg kg-1, p.o., SAL+ISCH group (n=6)] for 2 weeks after the induction of ischemia. The black ginseng and saline were administered orally everyday in the morning. The naïve normal group (Normal, n=6) was not treated with any drug and operation. The water maze tests were performed the last (second) week.
Morris water maze test
The swimming pool of the Morris water maze was a circular water tank 200 cm in diameter and 35 cm deep. It was filled to a depth of 21 cm with water at 23±2℃. A platform 15 cm in diameter and 20 cm in height was placed inside the tank with its top surface being 1.5 cm below the surface of the water. The pool was surrounded by many cues that were external to the maze [18]. A CCD camera was equipped with a personal computer for the behavioral analysis. Each rat was received four daily trials. For 6 consecutive days, the rats were tested with three acquisition tests. For the acquisition test, the rat was allowed to search for the hidden platform for 180 s and the latency to escape onto the platform was recorded. The animals were trained to find the platform that was in a fixed position during 6 days for the acquisition test. The inter-trial interval time was 1 min. The performance of the test animals in each water maze trial was assessed by a personal computer for the behavioral analysis (S-mart program, Spain).
Cresyl violet staining
Briefly, the rats were anesthetized (sodium pentobarbital, 100 mg/kg, ip), they were perfused transcardially with heparinized phosphate-buffered saline (PBS; pH 7.4) for 30 s and this was followed by 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4) for 10~15 min. The brains were postfixed in the same fixative overnight, cryoprotected in 30% sucrose solution in PBS, embedded and serially sectioned on a cryostat (Leica, Germany) at 30µm thickness in the coronal plane and then they were collected in PBS. One set of sections that represented different region of the cortex was dehydrated, rehydrated and stained with cresyl violet (ICN Biomedicals, Aurora, USA) to assess the cell loss and the morphology of the hippocampal area.
NADPH-d staining
The sections were stained for NADPH-d activity as previously described [19,20]. In brief, free-floating sections were incubated at 37℃ for 30 min in 0.05 M PBS containing 0.3% Triton X-100, 0.1 mg/ml nitroblue tetrazolium (Sigma, MO, USA) and 0.1 mg/ml β-NADPH (Sigma). The sections were washed three times with 0.05 M PBS and then they were mounted on gelatin-coated slides. The slides were air-dried overnight at room temperature, rinsed twice with distilled water and dried again. Coverslips were mounted using permount solution. The brain sections were analyzed using the atlas by Paxinos and Watson. The staining intensities of the sections that stained specifically for NADPH-d were quantitatively assessed according to a microdensitometrical method based on optical density and using an image analyzer (Multiscan, Fullerton, CA, USA) [21].
ChAT immunohistochemistry
The primary antibodies against the following specific antigen were used: cholineacetyl transferase (sheep polyclonal ChAT, concentration 1:2,000; Cambridge Research Biochemicals, Wilmington, D.E.). The primary antibody was prepared at a dilution in 0.3% PBST, 2% blocking serum and 0.001% kehole limpit hemocyanin (Sigma, USA). The sections were incubated in the primary antiserum for 72 h at 4℃. After three more rinses in PBST, the sections were placed in Vectastain Elite ABC reagent (Vector laboratories, Burlingame, CA) for 2 h at room temperature. Following a further rinsing in PBS, the tissue was developed using diaminobenzadine (Sigma, USA) as the chromogen. Images were captured using a DP2-BSW imaging system (Olympus, CA, USA) and they were processed using Adobe Photoshop. For measuring cells that were immunoreactive for ChAT, a grid was placed on the CA1 and CA3 areas of the hippocampus according to the method of Paxinos et al. [21]. The number of cells was counted at 100× magnification using a microscope rectangle grid that measured 200×200 mm2. The cells were counted in three sections per rat within the hippocampal CA1 and CA3 areas.
Statistical analysis
Statistical comparisons were done for the behavioral and histochemical studies using one-way ANOVA and repeated measures of ANOVA, respectively, and Tukey post hoc was done. All of the results are presented as means±S.E.M., and we used SPSS 15.0 for Windows for analysis of the statistics. The level of significance was set at p<0.05.
Go to :
RESULTS
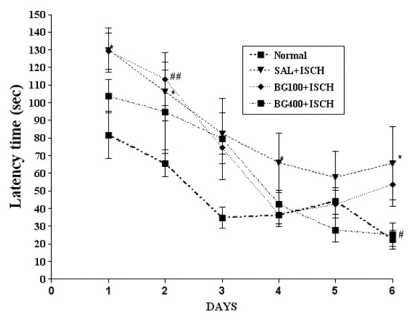
Effect of BG on the performance in the water maze test The effect of BG (100 or 400 mg/kg) on spatial learning was evaluated in the Morris water maze test. As shown in Fig. 1, the escape latency of the SAL+ISCH group was longer due to memory impairment than that of the normal group during all the trial sessions [p<0.05]. Data shows the mean group latencies to reach to hidden platform in the MWM for all the groups for 4 days. The escape latency differed among the groups when the results were averaged over all the sessions. The SAL+ISCH group showed a worse performance than did the normal group (at the 2nd day (p=0.004), 3rd day (p=0.022) and 4th day (p=0.05)). However, BG treatment had no significant interaction effect on the distance traveled to reach the platform from the 6th day.
Cresyl violet positive neurons of the hippocampus
The results of the evaluations of the cresyl violet positive neurons per section, from different hippocampal formations, are shown in Fig. 2. Post-hoc comparisons indicated that the cresyl violet activity in the hippocampus of the SAL+ISCH group was significantly lower than that of the normal group (p<0.05). In particular, there were significant differences in both the CA1 (F3,37=17.2, p=0.001) and CA3 (F3,37=10.6, p=0.001) areas. However, the number of cresyl violet positive neurons in the BG100+ISCH group was higher than that of the SAL+ISCH group, and particularly in the CA1 (p=0.001) and CA3 (p=0.001) areas. However, the number of cresyl violet positive neurons in the hippocampus of the BG100+ISCH group was not significantly different compared to that of the SAL+ISCH group, but there was a slight trend for a significant interaction effect on the expression of the cresyl violet positive neurons in the hippocampus. We also found that the tissue damage of the cortex was significantly decreased in the BG treated groups compared to that of the SAL+ISCH group.
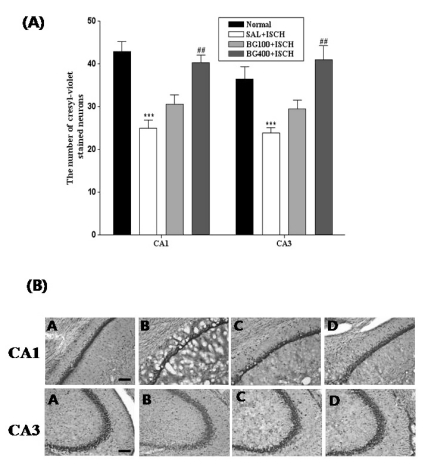 | Fig. 2(A) The number of cresyl violet stained neurons in the different hippocampal CA1,CA3 and DG areas of the experimental groups. Each value represents the mean±S.E.M. ***p<0.001 vs. the normal group and ##p<0.01 vs. the SAL+ISCH group. (B) Photographs showing the distribution of cresyl violet stained neurons in the hippocampus of the Normal (A), SAL+ISCH (B), BG100+ISCH (C) and BG400+ISCH (D). Sections were cut coronally at 30µm and the scale bar represents 200µm.
|
NADPH-d positive neurons of the hippocampus
The results of the evaluations of the NADPH-d-positive neurons per section, from different hippocampal formations, are shown in Fig. 3. Post-hoc comparisons indicated that the number of NADPH-d-positive neurons in the hippocampus of the SAL+ISCH group were noticeably less than that of the normal group (p<0.001). In particular, there were significant differences in both the CA3 area (F3,19=13.6, p=0.001) and the dentate gyrus (F3,19=7.1, p=0.003). However, the number of NADPH-d-positive neurons of the BG100+ISCH group was higher than that of the SAL+ISCH group, both in the CA3 area (p=0.001) and the DG (p=0.001).
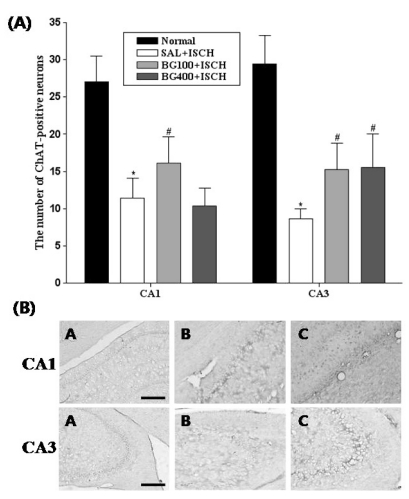 | Fig. 3(A) The number of choline acetyltransferase (ChAT) immunostained nuclei in the different hippocampal CA1 and CA3 areas of the experimental groups. Each value represents the mean±S.E.M. *p<0.05 vs. the normal group and #p<0.05 vs. the SAL+ISCH group. (B) Photographs showing the distribution of ChAT-immunoreactive cells in the hippocampus of the Normal (A), SAL+ISCH (B) and BG100+ISCH (C). Sections were cut coronally at 30µm and the scale bar represents 200µm.
|
ChAT immunoreactive neurons of the hippocampus
The results of the evaluations of the ChAT immunoreactive cells per section, from different hippocampal formations, are shown in Fig. 4. Post-hoc comparisons indicated that the ChAT activity in the hippocampus of the SAL+ISCH group was significantly lower than that of the normal group (p<0.05). In particular, there were significant differences in both the CA1 (F3,16=4.9, p=0.02) and CA3 (F3,16=9.3, p=0.001) areas. However, the ChAT reactivity in the BG treated groups was higher than that of the SAL+ISCH group, and particularly in the CA3 area (BG 100, p=0.05; BG 400, p=0.014). However, the ChAT reactivity in the hippocampal CA1 area showed no significant differences among the groups.
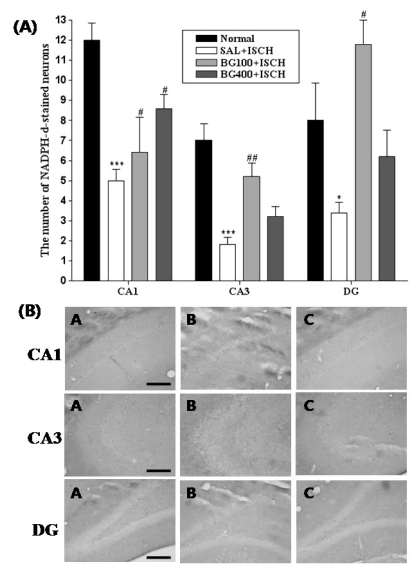 | Fig. 4(A) The number of NADPH-d positive neurons in the different hippocampal CA1 and CA3 areas of the experimental groups. Each value represents the mean±S.E.M. *p<0.05, ***p<0.001 vs. the normal group and #p<0.05, ##p<0.01 vs. the SAL+ISCH group. (B) Photographs showing the distribution of NADPH-d positive neurons in the hippocampus of the SHAM (A), SAL+ISCH (B), BG100+ISCH (C). Sections were cut coronally at 30µm and the scale bar represents 200µm.
|
Go to :
DISCUSSION
In this study, we investigated whether treatment with BG can improve impairment of memory and affect the cholinergic cells in the hippocampus of ischemia-induced memory deficit rats. The results showed that BG ameliorated the measures of stress-induced deficits of memory on the MWM, and the BG treated rats displayed an increased number of cholinergic positive neurons in the hippocampal CA1 and CA3 areas of the rats.
The extent of brain damage produced by MCAO is known to be dependent on the degree of the ischemic insult and its duration. One of the most critical factors is the period of occlusion. The damage produced by mild injury (30 min of MCAO) was confined to the striatum or cortex, whereas more than 2 h exposure to MCAO led to damage of the striatum, cortex and more remote areas, including the hippocampus [22]. In this study, the Morris water maze is well-established paradigm for evaluating deficits in hippocampal-dependent memory, and the MWM spatial learning task has been used for the validation of rodent models for neurocognitive disorders and for the evaluation of possible neurocognitive treatments [18,23-27]. In this study, the spatial memory tended to be improved in BG400 group during the training day (on the day 6), but not in the BG100 group. The data of the memory test demonstrated that BG protects the animals from the ischemia-induced decrease of the spatial learning and memory.
The neuroprotective effects of these natural drugs on the central acetylcholine system were also examined by the histochemistry of the hippocampal neurons. The degeneration of the cholinergic innervation from the basal forebrain to the hippocampal formation in the temporal lobe is thought to be one of the factors determining the progression of memory decay, both during normal aging and AD [28]. The best available marker for cholinergic neurons in the basal forebrain is ChAT activity. ChAT synthesizes the neurotransmitter acetylcholine in the basal forebrain and the cortex, hippocampus and amygdala. A significant reduction in ChAT activity in the postmortem brains of demented patients has been reported. In addition, there was a 20~50% decrease in ChAT activity in the hippocampus of the memory deficit rats. However, the present results show that BG exerts beneficial effects on cholinergic neurotransmission in the brain by increasing the hippocampal CA3 ChAT activities. However, our results showed that the levels of AchE in the hippocampus were not significantly different among all the groups.
NO-mediated mechanisms have been ascribed a role in cortical perfusion [29], learning and memory [30], as well as in neuronal plasticity [31,32]. Dysfunction of the neocortical and hippocampal cholinergic transmission has also been associated with reduced cortical cerebral blood flow and impaired learning and memory [33-35], suggesting a coupling of the basal forebrain cholinergic system. NADPH-diaphorase (NADPH-d) has been characterized biochemically and immunochemically as nitric oxide synthase [36-38]. Faber-Zuschratter et al. [39] demonstrated by ultrastructural studies that NADPH-d histochemical activity is located at both the pre- and postsynaptic membranes after long term potentiation (LTP). These mechanisms seem to be dependent on nitric oxide (NO) in the striatum, cerebellum and hippocampus [40-45]. Also, Chien et al. [46] reported that a NO synthesis inhibitor induced impaired learning and memory. Also, there is a specific decline of NOS-containing neurons in the hippocampal subregions CA1 and CA3 n brain form Alzheimer patients [47]. In agreement with the previous study, we found significant increased numbers of NADPH-d positive neurons in the hippocampal CA1 and CA3 areas and the dentate gyrus of the BG treated groups.
In summary, treatment with BG attenuated the ischemia-induced learning and memory deficits on the Morris water maze and treatment with BG had a protective effect against the ischemia-induced decreased number of cholinergic neurons. Also, treatment with BG increased NO synthesis in the hippocampus. BG is a good candidate for further investigations that may ultimately result in its clinical use. Further research that will examine the effects of BG activation on additional behavioral tasks will help to elucidate whether increasing the number of cholinergic neurons and nitric oxide synthesis may also facilitate other types of memory.
Go to :
 XML Download
XML Download