

 PDF
PDF ePub
ePub Citation
Citation Print
Print


ABBREVIATIONS
ROS
reactive oxygen species
IL
interleukin
NADPH
nicotinamide adenine dinucleotide phosphate (reduced form)
MAPK
mitogen-activated protein kinase
ERK
extracellular signal-regulated kinases
PI3K
phosphatidyl inositol 3-kinase
TNF
tumor necrosis factor
HPLC
high performance liquid chromatography
DMSO
dimethyl sulfoxide
MTT
3-(4, 5-dimethylthiozol-2-yl)-2,5-diphenyltetrazolium bromide
GAPDH
Glyceraldehyde-3-phosphate dehydrogenase
INTRODUCTION
Ethanol is a commonly used and abused substance. Notably, 10~13% ethanol is used in beverage such as wines [1]. Alcohol has diverse effects on human health and organ systems. Notably, ethanol intake injures the functional and structural integrity of the intestinal mucosa [2]. Clinical experience suggests that the frequency of chronic esophagitis is increased in patients who abuse alcohol [3]. Both the induction of gastro-esophageal reflux and disordered motility of the esophagus caused by acute ethanol ingestion may promote the development of mucosal lesions [3-5].
Reactive oxygen species (ROS) participate and regulate diverse downstream signaling pathways leading to specific cellular functions [6,7] such as growth, metabolic rate, cell division, necrosis, apoptosis and aging processes [8,9]. Oxygen-derived free radicals have been known to play a key role in the generation of gastrointestinal diseases, including the acid-related peptic diseases and inflammatory disorders [10,11]. It was demonstrated that cell damage caused by free radicals in gastric or esophageal mucosa can be prevented by the administration of free radical scavengers [12-14].
Many cellular responses to ethanol are mediated by the modulation of mitogen activated protein kinases (MAPK) signaling [15,16]. MAPK signaling cascades regulate important cellular processes including gene expression, cell proliferation, cell survival and death, and cell motility [17] and the induction of most cytokine genes requires activation of the ERK and p38 MAPK in response to a variety of extracellular stimuli [18,19]. ERK has been classically associated with growth and differentiation inducing signals, whereas p38 MAPK is involved in inflammatory cytokines and environmental stress inducers [20]. Several studies in human and animal models have shown that prolonged alcohol consumption is associated with elevated serum levels of not only tumor necrosis factor (TNF), but also interleukins such as IL-1, IL-6, and IL-8 [21-25]. Enhanced production of IL-1β and IL-6 has been documented in esophageal tissue of cats with experimental esophagitis [26,27], and both cytokines contribute to reduce esophageal circular muscle contractility [28].
Flavonoids are a large heterogeneous group of benzo-γ-pyron derivatives which are present in fruits, vegetables and medicinal herbs. Flavonoids have received a great deal of attention over the last several decades, and several biological activities including antioxidant, apoptosis-induction and anti-inflammatory effects have been identified [29,30]. Plant-originated flavonoids are highly gastroprotective against gastric mucosal lesions induced by ethanol in rats in vivo [31]. Among them, quercetin (3,5,7,3'4'-pentahydroxy flavon) has been known to possess a broad range of pharmacological properties, including anti-inflammatory [32], antioxidative [33] and anti-proliferative activities [34]. It has been reported that quercetin exerts potential gastroprotective effect against ethanol-induced cellular damages [35] and prevents the HCl- as well as ethanol-induced gastric mucosal injuries [36]. Quercetin-3-O-β-D-glucuronopyranoside (QGC) is a flavonoid glucoside extracted from Rumex Aquaticus Herba. In our previous study, QGC was more potent than quercetin on inhibition of experimental reflux esophagitis and indomethacin-induced gastritis in rats [37]. In another previous study, QGC enhanced antioxidant enzyme defense systems via heme oxygenase-1 (HO-1) expression and NF-E2-related factor 2 (Nrf2) translocation involving both ERK and PI3K-Akt pathways as well as PKC pathways in esophageal epithelial cell (EEC) [38]. In this study, we aimed to investigate the mechanism of the cytoprotective effect of QGC against cell damage induced by 10% ethanol in cultured feline EECs. As cell damage factors induced by 10% ethanol, we investigated intracellular ROS production, MAPK activation and IL-6 production and expression in this study.
Go to : 
METHODS
Materials
QGC (molecular weight: 477), which isolated from the herba of Rumex Aquaticus and its purity was 96~97%, was provided by Pharmacal Botany Resources Laboratory (Dr. Whang, Chung-Ang Univ., Seoul, Korea) and dissolved in serum free DMEM containing 0.01% dimethy sulfoxide (DMSO). Fetal bovine serum (FBS), antibiotic-antimycotic (penicillin, streptomycin, amphotericin B), trypsin-EDTA were purchased from Invitrogen (Grand Island, NY, USA); Dulbecco's modified Eagle's medium (DMEM), 0.1 N hydrochloric acid and phosphate-buffered saline (PBS) were from Welgene Inc. (Daegu, South Korea); absolute ethanol and DMSO were from Thermo Fisher Scientific (Waltham, MA, USA); actin and IL-6 antibodies were from Santa Cruz Biotechnology (Santa Cruz, CA, USA); goat anti-rabbit IgG-HRP and goat anti-mouse IgG-HRP were from Zymed Laboratories, Inc. (Eccles Avenue, CA, USA); phospho-specific ERK 1/2, phospho-JNKs and phospho-p38 MAPK were from Cell Signaling (Beverly, MA); Rainbow molecular weight marker were from Amersham (Arlington Heights, IL,USA); enhanced chemiluminescence (ECL) agents were from PerkinElmer Life Sciences (Boston, MA, USA); sodium dodecyl sulfate (SDS) sample buffer were from Owl Scientific, Inc. (Woburn, MA, USA); nitrocellulose membrane, Tris/Glycine/SDS buffer and Tris/Glycine buffer were from BioRad (Richmond, CA, USA); Restore™ Western Blot Stripping Buffer was from Pierce (Rockford, IL, USA); Hank's Balanced Salt Solution-Modified (HBSS), ammonium persulfate, ponceau S, bovine serum albumin (BSA), leupeptin, aprotinin, β-mercaptoethanol, N,N,N',N'-tetramethylethylene diamine (TEMED), ethylene glycolbis-(β-aminoethylether)-N,N,N',N'-tetraacetic acid (EGTA), ethylenediamine tetraacetic acid (EDTA), phenylmethylsulfonylfluoride (PMSF), 2',7'-Dichloro-fluorescein diacetate (DCF-DA) and other reagents were purchased from Sigma Chemical Co. (St. Louis, MO, USA);human IL-6 enzyme immunometric assay kit was from assay designs (Ann arbor, MI,USA).
Extraction and isolation of QGC from Rumex aquaticus herba
Fresh folium (600 g) of Rumex aquaticus (Korean name: Todaehwang) was extracted with ethanol under sonicator. After filtration, the ethanol solution was evaporated under a vacuum to yield an ethanol extract (72 g). The extract was partitioned between chlorform and water to give a chloroform-soluble fraction (16 g) and a water-soluble fraction (54 g). Based on target-guided fractionation, the watersoluble fraction was chromatographed on Sephadex LH 20 by elution with 50% methanol to give sub-fraction 1, 2, 3, 4. A portion of sub-fraction 2 was chromatographed with an ODS column using 30% methanol as eluent to give QGC (1.2 g) at a purity of 96~97% by HPLC.
QGC is yellow amorphous powder: IR (KBr) cm-1: 3385 (OH), 1657 (C=O), 1651, 1602, 1502 (aromatic ring), 1052 (glucuronide -CO); FAB-MS (neg.) m/z: 477[M-H]-, 301 [M-Glu]-; 1H-NMR and 13C-NMR data were consistent with literature values [39].
Preparation of cat esophageal epithelial tissue squares
All animal experiments were approved by the Institutional Animal Care and Use Committee of Chung-Ang University, in accordance with the guide for the Care and Use of Laboratory Animals in Seoul, Republic of Korea. Adult cats of either sex weighing between 2.5 and 4.0 kg were anesthetized with Zoletil 50 (12.5 mg/0.25 ml/kg), which composed of tiletamine and zolazepam, and euthanized with an overdose of 25% urethane (Aldrich, St. Louis, MO, USA). Then, the abdomen was opened with a midline incision and the esophagus was excised, cleaned and maintained in Krebs buffer with the following composition: 118 mM NaCl, 24 mM NaHCO3, 1.2 mM KH2PO4, 4.8 mM KCl, 2.5 mM CaCl2, 11 mM glucose and 1.2 mM MgSO4. The esophagus was opened along the lesser curvature. The location of the squamocolumnar junction was identified and the mucosa was peeled off. The submucosal connective tissue was then removed by micro spring scissors. The mucosa from the esophagus was sliced off into 0.5 mm thick sections with a Stadie Riggs tissue slicer (Thomas Scientific Apparatus, Philadelphia, PA, USA). The last slices were cut into 2×2 mm tissue squares with scissors.
Primary culture and identification of EECs
EECs were successfully isolated from mucosa and subcultured as previously described [40]. Briefly, the sliced tissue was then placed into DMEM supplemented with 10% FBS containing 100 U/ml penicillin, 0.1 mg/ml streptomycin and 0.25 µg/ml amphotericin B, then incubated in a humidified atmosphere of 5% CO2 and 95% air at 37℃. After 10 days, the medium was exchanged with fresh DMEM containing 10% FBS. After reaching confluence, cells were detached with 1% Trypsin-EDTA in HBSS with bicarbonate. Cells were then counted, seeded at 2×105 cells/ml on 100-mm culture dishes and maintained in DMEM containing 10% FBS. The medium was changed every 48 h until the cells reached confluence. Experiments were performed on cells at passage 3 or 4.
In order to confirm typical epithelial morphology of the primary-cultured EECs, cells were captured and confirmed using a phase contrast microscope (model ULWCD 0.30 Olympus, Tokyo, Japan) and a digital closed-circuit video camera (CCD color camera, Toshiba, Tokyo, Japan) connected to a Macintosh computer (Apple, Cupertino, CA) with NIH Image 1.57 software (National Institutes of Health, Bethesda, MD). In order to characterize epithelial cells and to exclude contamination by smooth muscle cells and fibroblasts, cells were fixed with 10% formalin containing 0.1% Triton X-100 at passage 1 and 2, and identified by an indirect immunofluorescent staining method using a cytokeratin monoclonal antibody from Dako and captured and confirmed by a Olympus BX51 microscope.
Measurement of cell viability
Cell viability was determined by the conventional MTT reduction assay as previously described [40]. In this assay, viable cells convert MTT to insoluble blue formazan crystals by the mitochondrial respiratory chain enzyme succinate dehydrogenase. Cells were seeded at a density of 2×104/6-well plates and maintained in DMEM containing 10% FBS. Then, cells were made quiescent at confluence by incubation in serum-free DMEM for 24 h, followed by treatment with indicated concentrations of each compound for the desired time. After the incubation, cells were washed with PBS three times and treated with MTT solution (final concentration, 5 mg/ml) for 4 h at 37℃. Then, the supernatant was removed and the formazan crystals were dissolved with 500 µl DMSO. Absorbance at 570 nm was measured with a microplate reader (Molecular Devices, Sunnyvale, CA).
Measurement of ROS production
2',7'-Dichlorofluorescein diacetate (DCF-DA) was used to measure the level of intracellular H2O2 production induced by ethanol in EECs [40]. Cells were seeded and grown on 6-well plates for 2 day and then serum-starved in DMEM for 24 h. The cells were treated with 50 µM QGC for 3 h and then incubated with H2O2-sensitive fluorophore DCFDA (20 µM) for 1 h at 37℃ in the dark. The cells were then washed with Krebs solution and exposed with ethanol for indicated times at 37℃ in the dark. The cells were harvested and centrifuged for 5 min at 10,000 rpm at 4℃ to remove the supernatants. After the pellets were resuspended with Krebs buffer, DCF fluorescence was measured using a fluorospectrophotometer using excitation and emission wavelengths of 485 and 535 nm, respectively (Tecan, GENios Pro).
Measurement of IL-6 production
Cells were cultured in 24-well plates until confluent and then treated with 50 µM QGC for 12 h before incubation with 10% ethanol for indicated times. After the incubation, supernatants were collected at various time points, and cytokine content was measured by an IL-6 enzyme immunometric assay kit. Assays were performed according to the manufacturer's instruction.
Preparation of cell extracts and protein determination
When the cells reached confluence, they were serum starved by incubation in serum-free DMEM for 24 h. The cells were then stimulated with each compound for indicated time periods or at indicated concentrations. After incubation, the cells were rapidly washed twice with icecold PBS and lysed with ice-cold lysis buffer for 5 min containing: 20 mM Tris- HCl (pH 7.4), 0.5 mM EDTA, 0.5 mM EGTA, 1% (w/v) Triton X-100, 0.01% (w/v) SDS, 10 µg/ml leupeptin, 10 µg/ml aprotinin, 1 mM phenylmethylsulfonyl fluoride, and 0.7 µg/ml β-mercaptoethanol. The lysates were scraped with a cell scraper and collected in eppendorf tubes. They were then sonicated and centrifuged for 10 min at 13,000 rpm at 4℃ to remove cellular debris and the supernatants were collected.
Equal amounts of the protein from each sample were resolved on a SDS-polyacrylamide gel by electrophoresis; the protein concentrations of supernatants were determined with Bradford reagent according to the instructions of the manufacturer (Bio-Rad Chemical Division, Richmond, CA). Absorption was monitored at 595 nm.
Western blotting analysis
Equal amounts of protein from each sample were subjected to electrophoresis on a 10% SDS-polyacrylamide gel and transferred to a nitrocellulose (NC) membrane, using a Power Pac 1000 power supply (Bio-Rad, Melville, NY). To block nonspecific binding, the NC membrane was incubated in 5% nonfat dry milk in PBS for 60 min followed by three rinses in milk-free PBS. The membranes were incubated for 1 h with shaking with primary antibodies raised against each phospho-specific ERK 1/2, phospho-JNKs, phospho-p38 MAPK and IL-6 protein followed by three washes with PBS containing 0.05% Tween 20. This was followed by 60 min incubation in horseradish peroxidase-conjugated secondary antibody. Detection was performed with an enhanced chemiluminescence agent. Molecular masses were estimated by comparison with a prestained molecular mass marker. To confirm uniformity of protein loading, the same blots were subsequently stripped with western blot stripping buffer and reprobed with ERK 1/2, p38 MAPK, JNK, and GAPDH antibody. Developed films were scanned and analyzed densitometrically using Scion Image. Percent of MAPKs activation and IL-6 expression were calculated as the ratio of phosphorylated MAPK to total MAPK and IL-6 to actin, respectively.
Statistical analysis
Data are expressed as means±SEM of n separate experiments and differences between means were analyzed by a Student's t test (two-tailed), with p<0.05 considered as significant.
Go to :
RESULTS
Time-course analysis for the effect of 10% ethanol on cell viability
To assess whether 10% ethanol causes cell death in cultured EECs, serum starved cells were exposed to 10% ethanol for the indicated times and then cell viability was measured using MTT assay (Fig. 1A). There was no significant decrease in cell viability of cultured EECs incubated in ethanol for 10 min compared to control. After exposure to ethanol for 10 min, however, cell viability was decreased to about 70%. In addition, exposure of cells to ethanol for 30 min led to less than 50% cell viability.
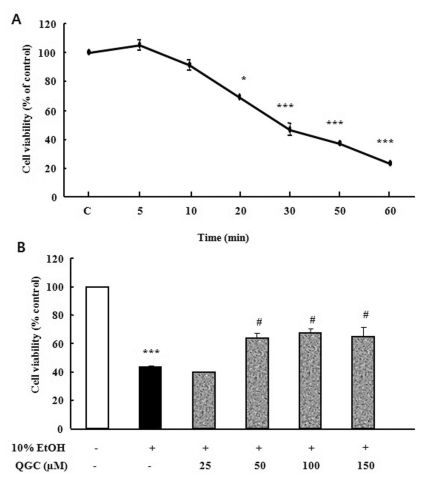 | Fig. 1Time course analysis for the effect of 10% ethanol in the absence or presence of QGC on the viability of cultured EECs. (A) Serum-starved EECs were stimulated with ethanol for the indicated times and their survival was estimated using the MTT assay. (B) Serum-starved EECs were pre-incubated in the presence of QGC for 12 h at indicated concentrations. EECs were then stimulated with 10% ethanol for 30 min and their survival was estimated using MTT assay. Data are expressed as means±SEM of three experiments. Student's t-test; *p<0.05, ***p<0.001 vs. control, #p<0.05 vs. cells in 10% ethanol alone.
|
Concentration-related protective effect of QGC against the ethanol-induced cytotoxicity
In a previous study, we have reported the chemical structure of QGC compared to quercetin [38]. QGC alone did not exhibit any significant cytotoxicity on EECs at various concentration of 25~200 µM when incubated for 24 h (data not shown). In this time, to assess the cytoprotective effect of QGC against 10% ethanol-induced cell damage, serum starved cells were pre-incubated with 25~150 µM QGC for 12 h and then exposed to 10% ethanol for 30 min, followed by measurement of cell viability (Fig. 1B). Ethanol treatment alone for 30 min caused a significant decrease to about 45% in cell viability. When cells were pretreated with 50~150 µM QGC for 12 h, the viability of cells exposed to ethanol significantly increased to 68%.
Effect of QGC on intracellular H2O2 levels
Serum starved EECs were pre-incubated with the presence of 50 µM QGC for 4 h. Cells were then stimulated with the 10% ethanol for 10 min and intracellular ROS production was estimated using DCF-DA (Fig. 2). 10% ethanol alone induced significant increase in intracellular ROS production by 135.3% vs. control. However, when cells were pretreated with 50 µM QGC for 4 h, the ROS levels were significantly decreased below about 45%.
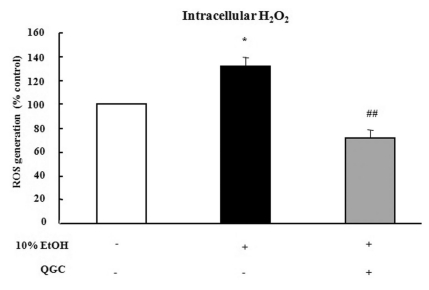 | Fig. 2Effect of QGC on intracellular H2O2 levels induced by ethanol in EECs. Serum-starved EECs were pre-incubated in the presence of 50 µM QGC for 4 h. EECs were then stimulated with ethanol for 10 min and ROS production was estimated using DCF-DA assay. Data are expressed as means±SEM of three experiments. Student's t-test; *p<0.05 vs. control, ##p<0.001 vs. cells in 10% ethanol alone.
|
Ethanol-induced MAPK activation
To test whether ethanol induces activation of MAPK, serum starved cells were treated with 10% ethanol at the indicated time periods. Ethanol induced the activation of ERK 1/2, which reached maximally at 10 min (Fig. 3A). Longer stimulation with 10% ethanol, only slightly reduced ERK 1/2 phosphorylation. As shown in Fig. 3B, phosphorylation of p38 MAPK did not cause significant changes after treatment of ethanol. Similarly, JNK was also not changed (Fig. 3C).
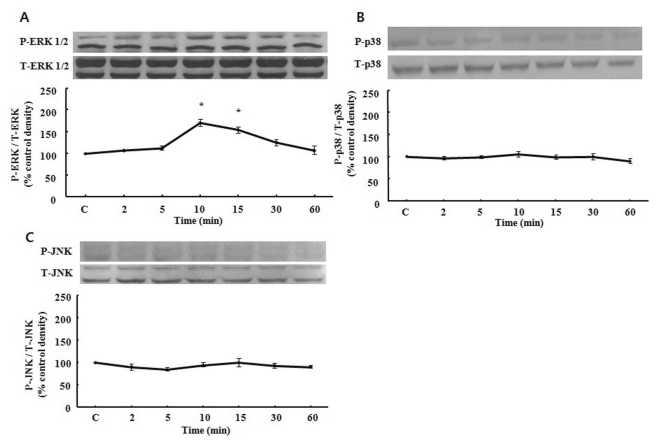 | Fig. 3Time course of MAPK phosphorylation induced by ethanol. Serum-starved EECs were incubated with ethanol for indicated time periods. Phosphorylation of ERK 1/2 (A), p38 MAPK (B) and JNK (C) were estimated by western blotting analysis. Data are expressed as means±SEM of three experiments. Student's t-test; *p<0.05 vs. control.
|
Effect of QGC and diphenylene iodonium (DPI) on ethanol-induced ERK 1/2 activation
Next, serum starved EECs were pre-incubated in the presence of 50 µM QGC for 12 h and then stimulated with 10% ethanol for 10 min. Pretreatment with QGC inhibited the expression of 10% ethanol-induced ERK 1/2 phosphorylation. To further clarify whether intracellular ROS production is involved in 10% ethanol-induced ERK 1/2 activation, DPI was utilized. Serum starved cells were pretreated with 10 µM DPI for 30 min and then exposed to 10% ethanol for 10 min. DPI blocked ethanol-induced activation of ERK 1/2 (Fig. 4). These results indicated that the inhibitory effect of QGC on ERK 1/2 activation by ethanol treatment may be mediated by reducing ethanol-induced intracellular ROS production which originate from NADPH oxidase.
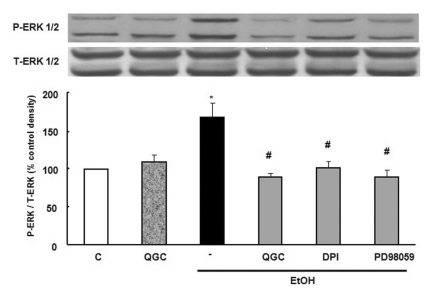 | Fig. 4Effects of QGC and DPI on ethanol-induced ERK 1/2 phosphorylation. Serum-starved EECs were incubated with QGC (50 µM, 12 h), DPI (a NADPH oxidase inhibitor, 10 µM, 0.5 h), PD98059 (a MEK inhibitor, 30 µM, 1 h) prior to ethanol treatment for 10 min. Phosphorylation of ERK 1/2 was estimated by western blot analysis. Data are expressed as means±SEM of three experiments.
Student's t-test; *p<0.05 vs. control, #p<0.05 vs. cells in 10% ethanol alone.
|
Effect of ethanol in the presence or absence of QGC on IL-6 production and expression
To test whether ethanol induces a cytokine response, serum starved EECs were stimulated with 10% ethanol for 30 and 60 min. Ethanol alone did not induce IL-6 protein expression in the absence or presence of QGC when estimated by western blotting (Fig. 5A). Moreover, ethanol also did not induce IL-6 release in the culture medium in the absence or presence of QGC compared to control (Fig. 5B).
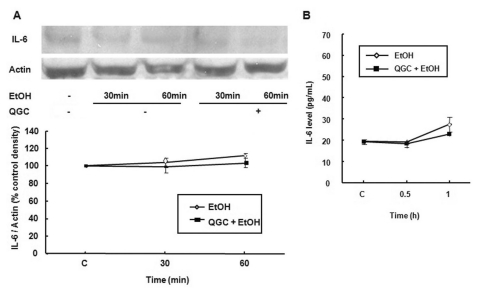 | Fig. 5Effect of ethanol in the absence or presence of QGC on IL-6 protein expression or release in EECs. Serum-starved EECs were stimulated with ethanol for 30 and 60 min in the absence or presence of 50 µM QGC for 12 h. (A) IL-6 expression was estimated by western blot analysis. (B) IL-6 production in culture medium was estimated by an EIA kit. Data are expressed as means±SEM of three experiments.
|
Go to :
DISCUSSION
Alcohol has many effects on the esophagus and stomach, and changes in these two organs can significantly increase morbidity due to alcohol consumption [3]. Ethanol is related to inflammation of the esophagus and stomach [41]. Both acute and chronic alcohol consumption have severe effects on the structure and function of the entire gastrointestinal tract which results in a vicious cycle [42]. Ethanol intake injures the functional and structural integrity of the intestinal mucosa [2]. Previous study showed that ethanol-induced cell injury was dependent on both the concentration of ethanol applied and the duration of exposure in rat gastric epithelial cells. 6~10% of ethanol was used as a cytotoxic agent [43]. In the present study, we also confirmed that 10% ethanol induces cytotoxicity to cat EECs.
It is well known that flavonoids, which are natural product of plants, have antioxidative and the antiinflammatory effects [44]. Quercetin has already been reported that antioxidative and antiinflammatory flavonoid [45,46]. In our previous study, QGC, a flavonoid glycoside extracted from Rumex Aquaticus, also acts as a non-stressful and non-cytotoxic antioxidant and antiinflammatory flavonoid in rat model [37]. Flavonoids are known as free radical scavengers and cytoprotective compounds [43], and exhibited protection against H2O2-mediated cytotoxicity. It was known that antioxidants protect cells against ethanol-induced cytotoxicity and apoptosis [47]. In our study, QGC also exhibited a protective effect against 10% ethanol which caused EEC death. We have already confirmed that QGC enhances antioxidant enzyme defense systems via HO-1 expression and Nrf2 translocation involving both the ERK and PI3K-Akt pathways as well as partial involvement of PKC pathways in EEC [38]. Mice lacking functional HO-1 exhibit chronic inflammation and increased mortality after lipopolysaccharide challenge [48,49]. Overexpression of HO-1 in cells resulted in a marked reduction in injury and cytotoxicity induced by oxidative stress [50,51]. An HO-1 inhibitor, ZnPP significantly attenuated the cytoprotective effect of eupatilin, one of the pharmacologically active flavones derived from Artemisia plants, in indomethacin-induced cytotoxicity [52]. We presumed that the antioxidative effect mediated by QGC-induced HO-1 expression may contribute to cytoprotection of the ethanol-induced cytotoxicity. In addition, we measured directly the intracellular ROS production induced by ethanol in the absence or presence of QGC in order to confirm the antioxidative effect of QGC. Exposure to oxidant molecules from the environment, food or during pathologies can generate ROS. Overproduction of ROS can cause oxidative stress and disease [53]. Several studies have suggested that the generation of ROS may be involved in the pathogenesis of ethanol-induced gastric mucosal injury in vivo, or resulted in cell death of human colon cancer cell and neuronal cell [54-57]. Natural and synthetic antioxidants or induction of cellular antioxidant systems can modulate the adverse effects of oxidative stress [53]. Several previous studies have indicated that flavonoids exhibit potent down regulation of ROS generation [58,59]. In the present study, our results suggest that QGC acts as a scavenger of intracellular ROS generation induced by 10% ethanol in EECs.
Ethanol exposure causes the depletion of glutathione (GSH) and the formation of ROS [60,61]. It has been shown that ROS causes deleterious effects by oxidizing important structures in the cells [62] or acting as a second messenger stimulating intracellular signaling pathways including MAPK [63]. Modulation of MAPK signaling pathway by ethanol is distinctive, depending on the cell type, ethanol concentration and duration of exposure [15]. Acute exposure to ethanol results in activation of ERK in astrocytes. However, chronic ethanol treatment causes activation of ERK and p38 MAPK leading to increased synthesis of TNF [64]. Ethanol treatment of stromal osteoblasts increases the ROS associated with induction of NADPH oxidase (NOX) and downstream signaling cascades involving sustained activation of ERK [65]. Also, ROS produced by ethanol in liver Kupffer cells and stellate cells as well as in the lung occur, in part, through activation of NADPH oxidase [66-68]. Concordantly, our data also showed that ethanol induced ERK 1/2 activation via NADPH oxidase-derived ROS production and reduction of the ROS generation by QGC may contribute to inhibition of ethanol-induced ERK 1/2 activation in EECs.
Alcohol consumption is associated with elevated serum levels of interleukins such as IL-1, IL-6, and IL-8 [25,69]. Enhanced production of IL-1β and IL-6 has been documented in esophageal tissue of cats with experimental esophagitis. Mucosa from esophagitis patients has the highest concentrations of IL-1β and IL-6, cytokines known to reduce esophageal muscle contractility [28,70]. However, in our study, 10% ethanol in the absence or presence of QGC did not elicit any changes in the level of IL-6 protein expression or release. Although the ethanol did not induce any change of IL-6 level in our experimental condition, ethanol reduced cell viability (45% compared to control) and induced ERK 1/2 activation via intracellular H2O2 production. Conversely, co-treatment with QGC significantly reversed the reduction of ethanol-induced cell viability from 45% (ethanol alone) to 68% (ethanol with QGC). Moreover, QGC exhibited inhibitory effect on the ROS-dependent ERK 1/2 activation induced by ethanol. Although the mechanism underlying the effect of QGC was not detail determined within this study, these phenomena indicate that QGC has cytoprotective effect against ethanol-induced cytotoxicity. Just, the relationship between the cytoprotective effect by QGC and QGC-induced inhibitory effect on the ROS-dependent ERK 1/2 activation induced by ethanol should be explored via further study.
In conclusion, QGC reduce the 10% ethanol-induced cytotoxicity, and inhibits the production of intracellular ROS and ERK 1/2 activation induced by ethanol. Whether or not QGC-induced inhibitory effect on the ROS-dependent ERK 1/2 activation induced by ethanol is involved in the cytoprotective effect of QGC should be determined by further study.
Go to :
 XML Download
XML Download