

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Abstract
Golgi SNAP receptor complex 1 (GS28) has been implicated in vesicular transport between intra-Golgi networks and between endoplasmic reticulum (ER) and Golgi. Additional role(s) of GS28 within cells have not been well characterized. We observed decreased expression of GS28 in rat ischemic hippocampus. In this study, we examined the role of GS28 and its molecular mechanisms in neuronal (SK-N-SH) cell death induced by hydrogen peroxide (H2O2). GS28 siRNA-transfected cells treated with H2O2 showed a significant increase in cytotoxicity under glutathione (GSH)-depleted conditions after pretreatment with buthionine sulfoximine, which corresponded to an increase of intracellular reactive oxygen species (ROS) in the cells. Pretreatment of GS28 siRNA-transfected cells with p38 chemical inhibitor significantly inhibited cytotoxicity; we also observed that p38 was activated in the cells by immunoblot analysis. We confirmed the role of p38 MAPK in cotransfected cells with GS28 siRNA and p38 siRNA in the cell viability assay, flow cytometry, and immunoblot. Involvement of apoptotic or autophagic processes in the cells was not shown in the cell viability, flow cytometry, and immunoblot analyses. However, pretreatment of the cells with necrostatin-1 completely inhibited H2O2-induced cytotoxicity, ROS generation, and p38 activation, indicating that the cell death is necroptotic. Collectively these data imply that H2O2 induces necroptotic cell death in the GS28 siRNA-transfected cells and that the necroptotic signals are mediated by sequential activations in RIP1/p38/ROS. Taken together, these results indicate that GS28 has a protective role in H2O2-induced necroptosis via inhibition of p38 MAPK in GSH-depleted neuronal cells.
Go to : 
References
1. Ryter SW, Kim HP, Hoetzel A, Park JW, Nakahira K, Wang X, Choi AM. Mechanisms of cell death in oxidative stress. Antioxid Redox Signal. 2007; 9:49–89.

2. Inoue M, Sato EF, Nishikawa M, Park AM, Kira Y, Imada I, Utsumi K. Mitochondrial generation of reactive oxygen species and its role in aerobic life. Curr Med Chem. 2003; 10:2495–2505.
3. Reiter RJ. Oxidative processes and antioxidative defense mechanisms in the aging brain. FASEB J. 1995; 9:526–533.
4. Schafer FQ, Buettner GR. Redox environment of the cell as viewed through the redox state of the glutathione disulfide/glutathione couple. Free Radic Biol Med. 2001; 30:1191–1212.
5. Martindale JL, Holbrook NJ. Cellular response to oxidative stress: signaling for suicide and survival. J Cell Physiol. 2002; 192:1–15.
6. Kim KC, Lee C. Curcumin induces downregulation of E2F4 expression and apoptotic cell death in HCT116 human colon cancer cells; involvement of reactive oxygen species. Korean J Physiol Pharmacol. 2010; 14:391–397.
7. Kitanaka C, Kuchino Y. Caspase-independent programmed cell death with necrotic morphology. Cell Death Differ. 1999; 6:508–515.
8. Degterev A, Huang Z, Boyce M, Li Y, Jagtap P, Mizushima N, Cuny GD, Mitchison TJ, Moskowitz MA, Yuan J. Chemical inhibitor of nonapoptotic cell death with therapeutic potential for ischemic brain injury. Nat Chem Biol. 2005; 1:112–119.
9. Azad MB, Chen Y, Gibson SB. Regulation of autophagy by reactive oxygen species (ROS): implications for cancer progression and treatment. Antioxid Redox Signal. 2009; 11:777–790.
10. Son MJ, Lee SB, Byun YJ, Lee HO, Kim HS, Kwon OJ, Jeong SW. Sodium nitroprusside induces autophagic cell death in glutathione-depleted osteoblasts. J Biochem Mol Toxicol. 2010; 24:313–322.
11. Jang H, Choi SY, Cho EJ, Youn HD. Cabin1 restrains p53 activity on chromatin. Nat Struct Mol Biol. 2009; 16:910–915.
12. Kim SY, Lee MY, Cho KC, Choi YS, Choi JS, Sung KW, Kwon OJ, Kim HS, Kim IK, Jeong SW. Alterations in mRNA expression of ribosomal protein S9 in hydrogen peroxide-treated neurotumor cells and in rat hippocampus after transient ischemia. Neurochem Res. 2003; 28:925–931.
13. Bains JS, Shaw CA. Neurodegenerative disorders in humans: the role of glutathione in oxidative stress-mediated neuronal death. Brain Res Brain Res Rev. 1997; 25:335–358.
14. Peuchen S, Bolaños JP, Heales SJ, Almeida A, Duchen MR, Clark JB. Interrelationships between astrocyte function, oxidative stress and antioxidant status within the central nervous system. Prog Neurobiol. 1997; 52:261–281.
15. Lang CA, Naryshkin S, Schneider DL, Mills BJ, Lindeman RD. Low blood glutathione levels in healthy aging adults. J Lab Clin Med. 1992; 120:720–725.
16. Byun YJ, Kim SK, Kim YM, Chae GT, Jeong SW, Lee SB. Hydrogen peroxide induces autophagic cell death in C6 glioma cells via BNIP3-mediated suppression of the mTOR pathway. Neurosci Lett. 2009; 461:131–135.
17. Zhang T, Hong W. Ykt6 forms a SNARE complex with syntaxin 5, GS28, and Bet1 and participates in a late stage in endoplasmic reticulum-Golgi transport. J Biol Chem. 2001; 276:27480–27487.
18. Maekawa M, Inoue T, Kobuna H, Nishimura T, Gengyo-Ando K, Mitani S, Arai H. Functional analysis of GS28, an intra-Golgi SNARE, in Caenorhabditis elegans. Genes Cells. 2009; 14:1003–1013.
19. Lewén A, Matz P, Chan PH. Free radical pathways in CNS injury. J Neurotrauma. 2000; 17:871–890.
20. Vandenabeele P, Galluzzi L, Vanden Berghe T, Kroemer G. Molecular mechanisms of necroptosis: an ordered cellular explosion. Nat Rev Mol Cell Biol. 2010; 11:700–714.
21. Rosenbaum DM, Degterev A, David J, Rosenbaum PS, Roth S, Grotta JC, Cuny GD, Yuan J, Savitz SI. Necroptosis, a novel form of caspase-independent cell death, contributes to neuronal damage in a retinal ischemia-reperfusion injury model. J Neurosci Res. 2010; 88:1569–1576.
22. Berghe TV, Vanlangenakker N, Parthoens E, Deckers W, Devos M, Festjens N, Guerin CJ, Brunk UT, Declercq W, Vandenabeele P. Necroptosis, necrosis and secondary necrosis converge on similar cellular disintegration features. Cell Death Differ. 2010; 17:922–930.
23. Ji D, Kamalden TA, del Olmo-Aguado S, Osborne NN. Light- and sodium azide-induced death of RGC-5 cells in culture occurs via different mechanisms. Apoptosis. 2011; 16:425–437.
24. Asare N, Låg M, Lagadic-Gossmann D, Rissel M, Schwarze P, Holme JA. 3-Nitrofluoranthene (3-NF) but not 3-aminofluoranthene (3-AF) elicits apoptosis as well as programmed necrosis in Hepa1c1c7 cells. Toxicology. 2009; 255:140–150.
Go to :
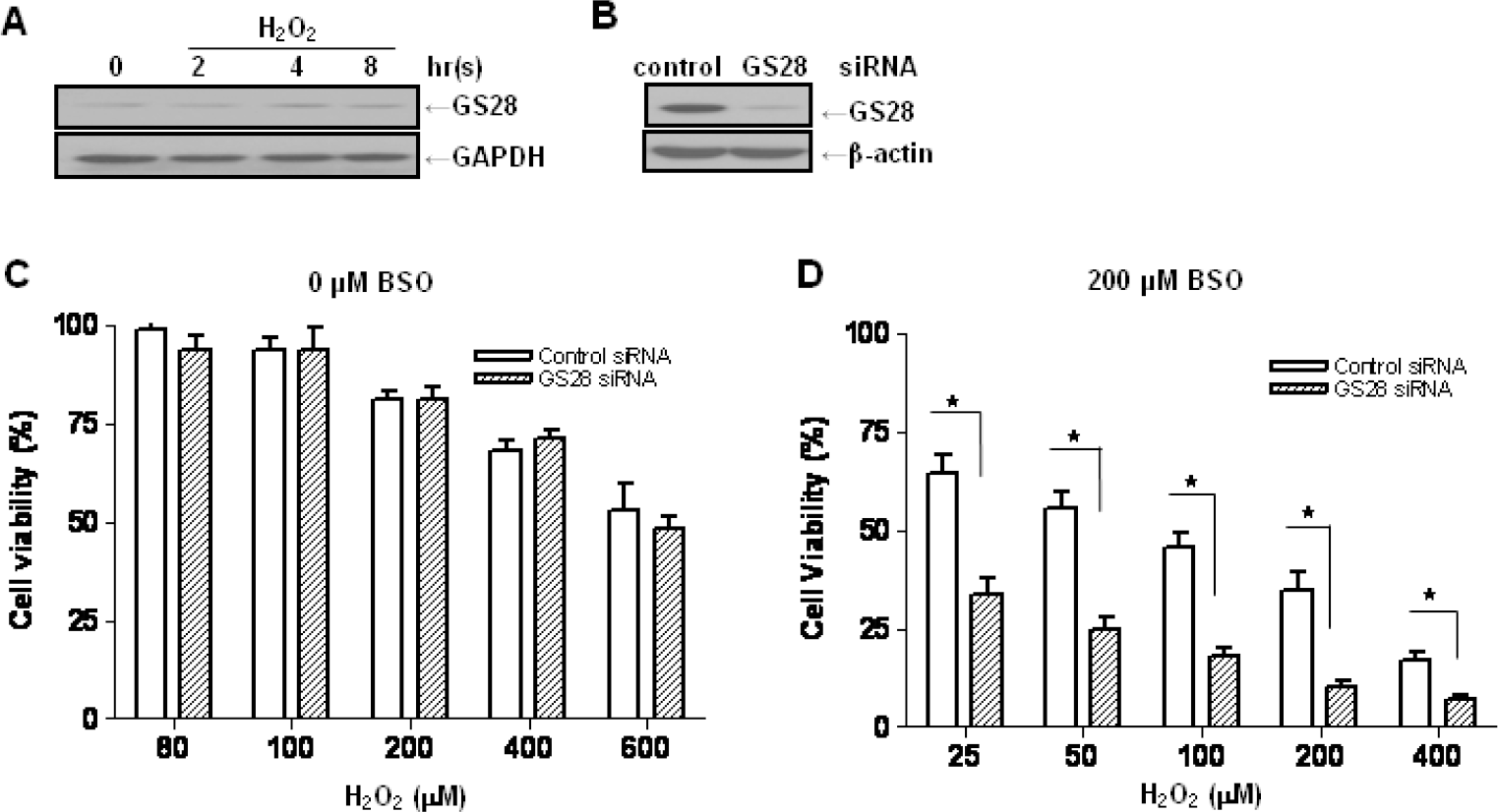 | Fig. 1.Increase of H2O2-induced cytotoxicity by silencing gene expression of the GS28 in glutathione (GSH)-depleted neuronal cells. (A) After treatment of neuroblastoma SK-N-SH cells with 400 μM H2O2 for indicated times, expression of GS28 in the cell lysates was examined by immunoblot analysis using GS28 or GAPDH antibody. (B) siRNAs were transfected into cells by lipofectamine, and efficiency of silencing GS28 expression was examined with immunoblot 3 days after transfection. The transfected cells were treated with various concentrations of H2O2 for 24 h in the absence (C) or presence (D) of 200 μM buthionine sulfoximine (BSO). Cell viability was determined by an MTT reduction assay. The data represent the mean±SEM of 3 independent experiments. ∗p<0.05 versus the values in control siRNA-transfected cells (two-way ANOVA, Bonferroni posttests). |
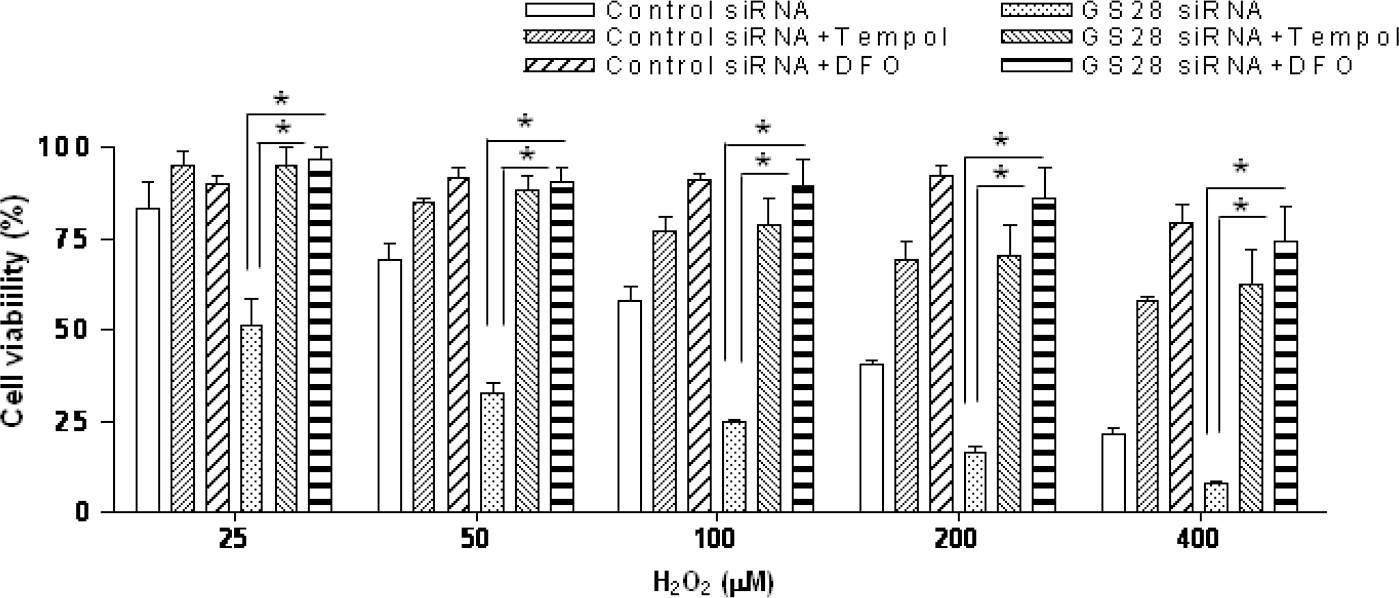 | Fig. 2.Contribution of H2O2-induced reactive oxygen species (ROS) to cytotoxicity in GS28 siRNA-transfected cells. Contribution of H2O2-induced ROS to cytotoxicity in GS28 siRNA-transfected cells was examined. siRNA-transfected cells were treated with H2O2 for 24 h in the presence of BSO without or with pretreatment with 1 mM tempol and 50 μM deferoxamine (DFO). Cell viability was determined by an MTT reduction assay. The data represent the mean±SEM of 3 independent experiments. ∗p<0.05 versus the values in GS28 siRNA-transfected cells without pretreatment. |
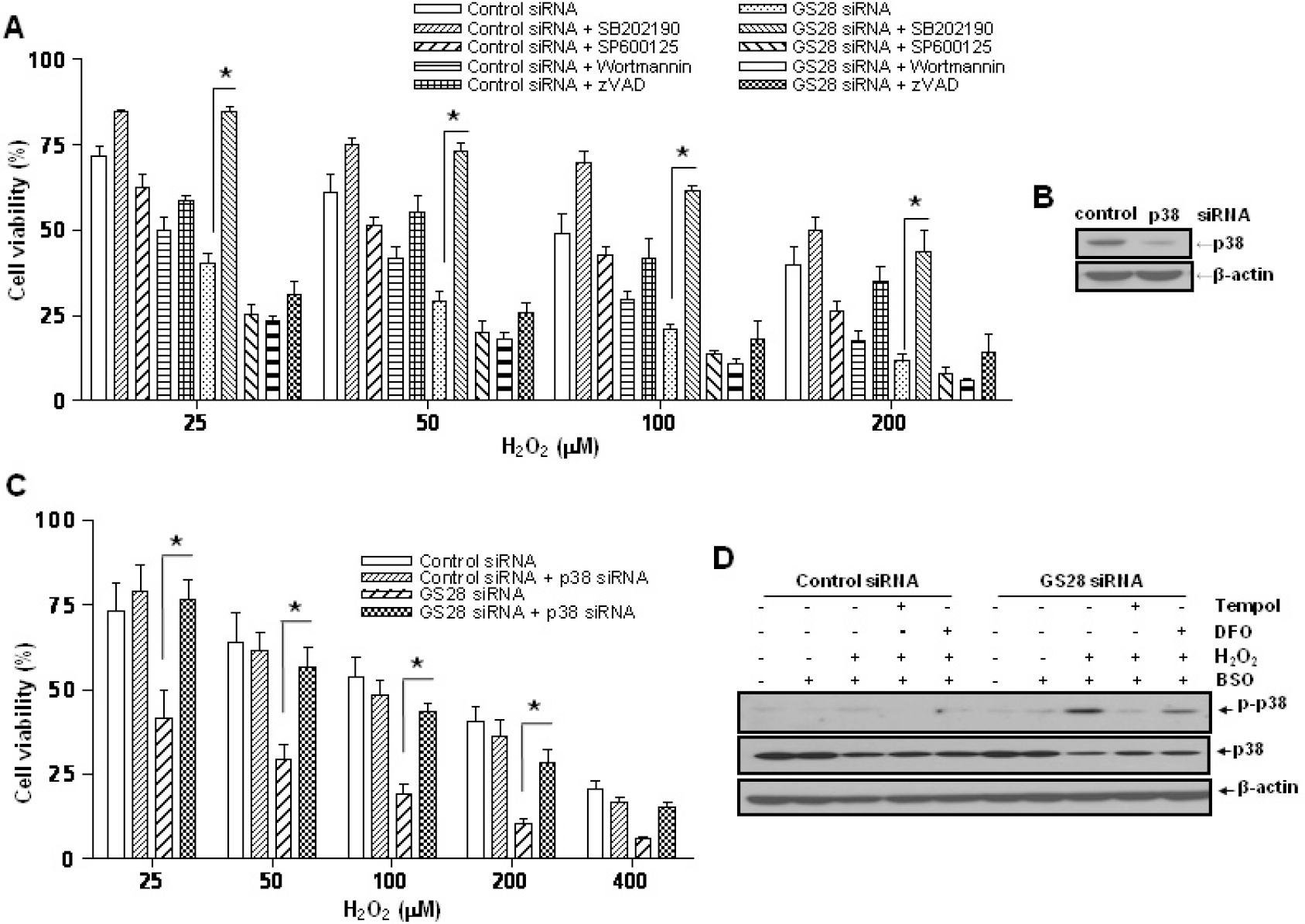 | Fig. 3.Involvement of p38 in the H2O2-induced cytotoxicity in GS28 siRNA-transfected cells. (A) siRNA-transfected cells were treated with H2O2 in the presence of BSO without or with pretreatment with chemical inhibitors. The inhibitors are 20 μM SB202190 (p38), 20 μM SP600125 (JNK), 200 nM wortmannin (PI3K), and 20 μM zVAD-fmk (pan-specific caspase). Cell viability was determined by an MTT reduction assay. The data represent the mean±SEM of 3 independent experiments. ∗p<0.05 versus the values in GS28 siRNA-transfected cells without pretreatment. (B) Efficiency of silencing gene expression was examined after transfection of p38 siRNA by immunoblot analysis of protein lysates. β-actin was used as loading controls. (C) H2O2-induced cytotoxicity was examined in GS28 siRNA-transfected cells without or with cointroduction of p38 siRNA. The cells were treated with H2O2 for 24 h in the presence of BSO. Cell viability was determined by an MTT reduction assay. The data represent the mean±SEM of 3 independent experiments. ∗p<0.05 versus the values in the cells transfected with GS28 siRNA alone. (D) Activation of p38 MAPK in GS28 siRNA-transfected cells treated with H2O2 was examined. siRNA-transfected cells were treated with 100 μM H2O2 for 3 h without or with pretreatment with 1 mM tempol and 50 μM deferoxamine (DFO). Phosphorylation of p38 was determined by immunoblot analysis. β-actin was used as a loading control. |
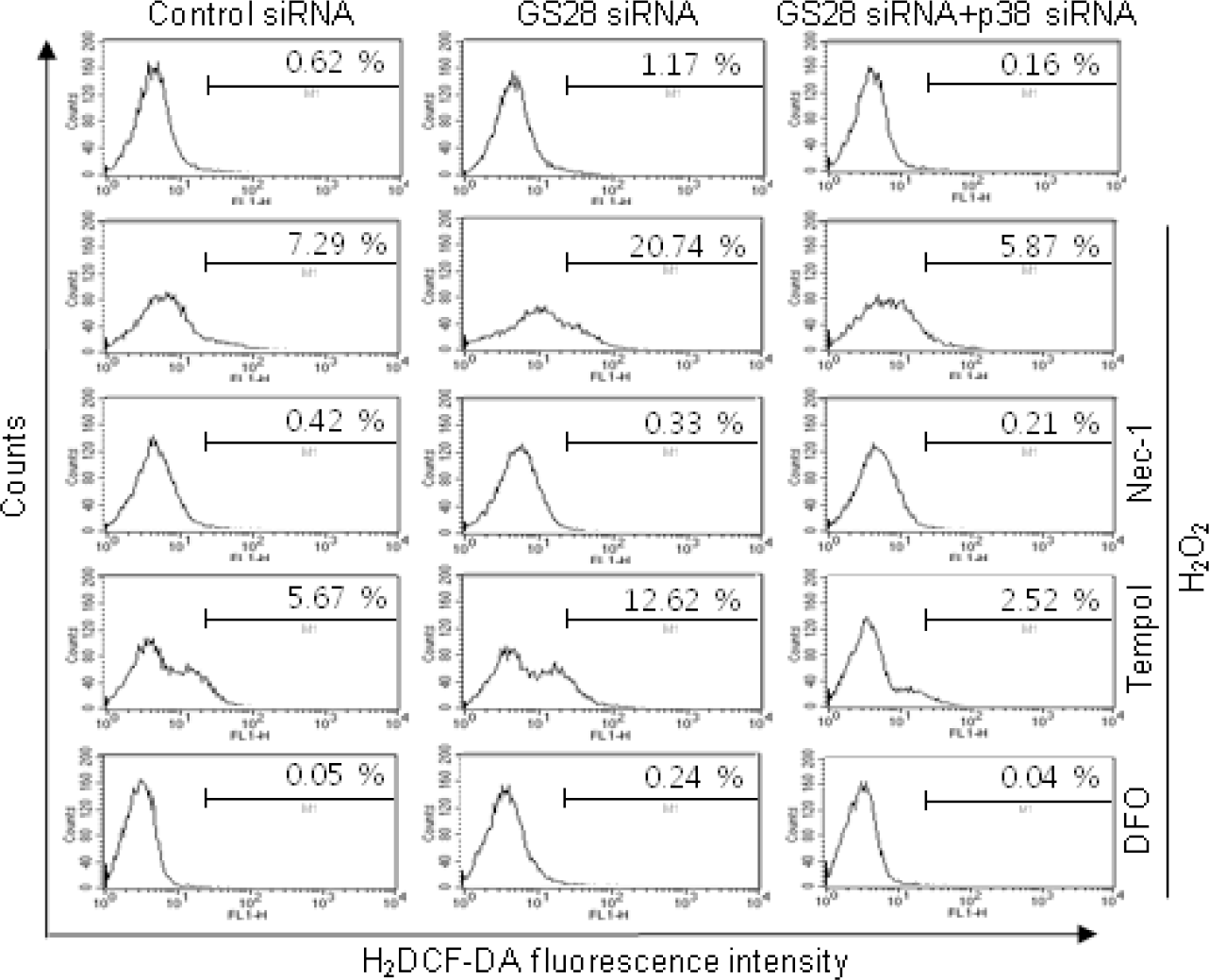 | Fig. 4.Determination of reactive oxygen species (ROS) generated by H2O2 in GS28 siRNA-transfected cells. siRNA-transfected cells were treated with 200 μM H2O2 for 3 h in the presence of BSO without or with pretreatment with 40 μM necrostatin-1 (Nec-1), 1 mM tempol and 50 μM deferoxamine (DFO). The cells were incubated with 5 μM H2DCF-DA, and fluorescence intensities were measured in flow cytometry, as described in Materials and Methods. |
 | Fig. 5.Immunoblot analysis in GS28 siRNA-transfected cells treated with H2O2. siRNA-transfected cells were treated with 100 μM H2O2 for indicated times. Activation of p38, procaspase-3, and LC3 and cleavage of PARP in the cells were examined by immunoblot analysis using the specific antibodies as described in Materials and Methods. β-actin was used as a loading control. |
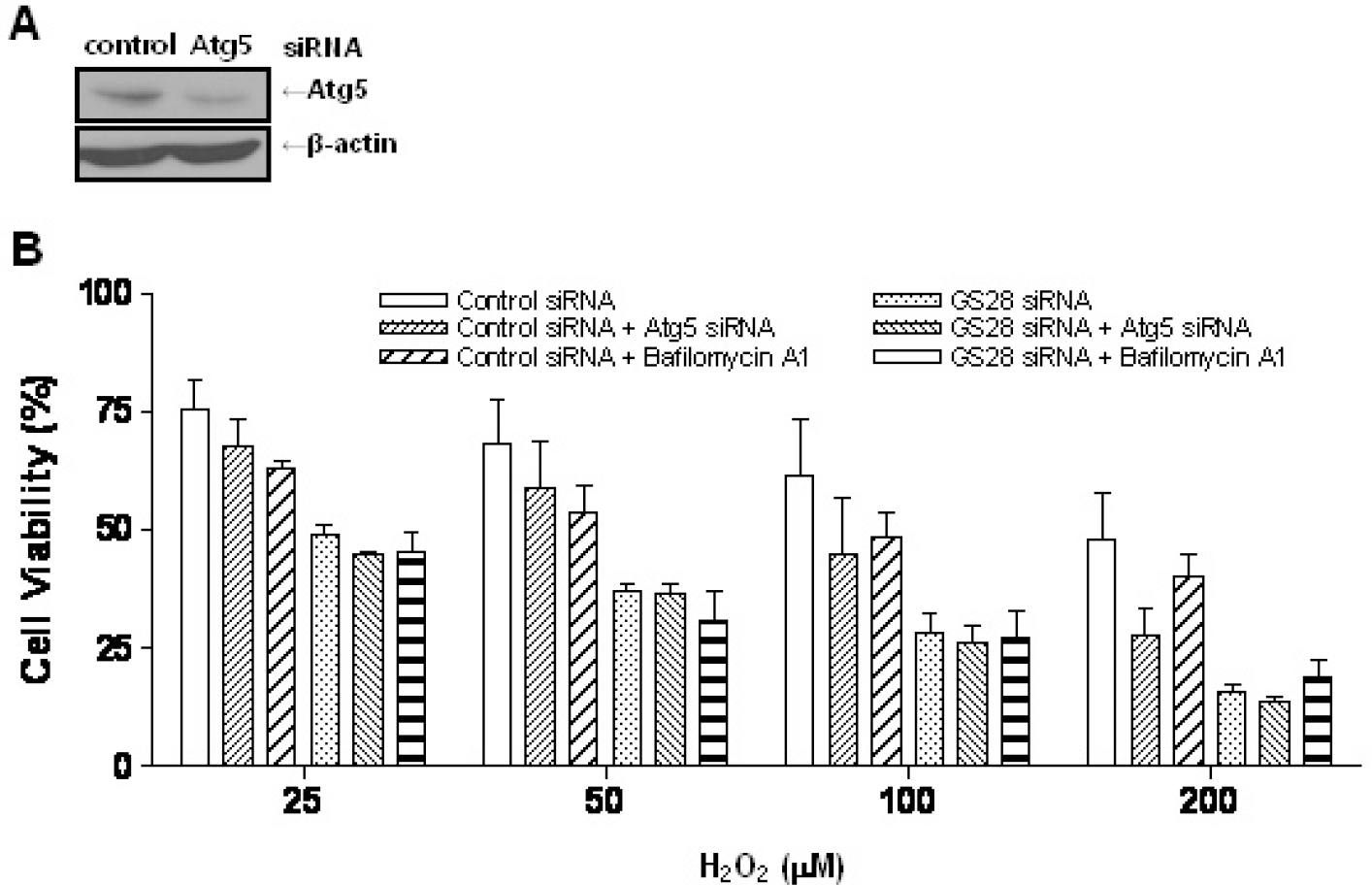 | Fig. 6.Non-autophagic cell death by H2O2 in GS28 siRNA-transfected cells. (A) Efficiency of silencing gene expression was examined after transfection of Atg5 siRNA by immunoblot analysis of protein lysates. β-actin was used as loading controls. (B) H2O2-induced cytotoxicity was examined in GS28 siRNA-transfected cells without or with cointroduction of Atg5 siRNA. The cells were treated with H2O2 for 24 h in the presence of BSO. Cell viability was determined by an MTT reduction assay. Effect of pretreatment with 50 nM bafilomycin A1 also was examined in GS28 siRNA-transfected cells treated with H2O2. The data represent the mean±SEM of 3 independent experiments. |
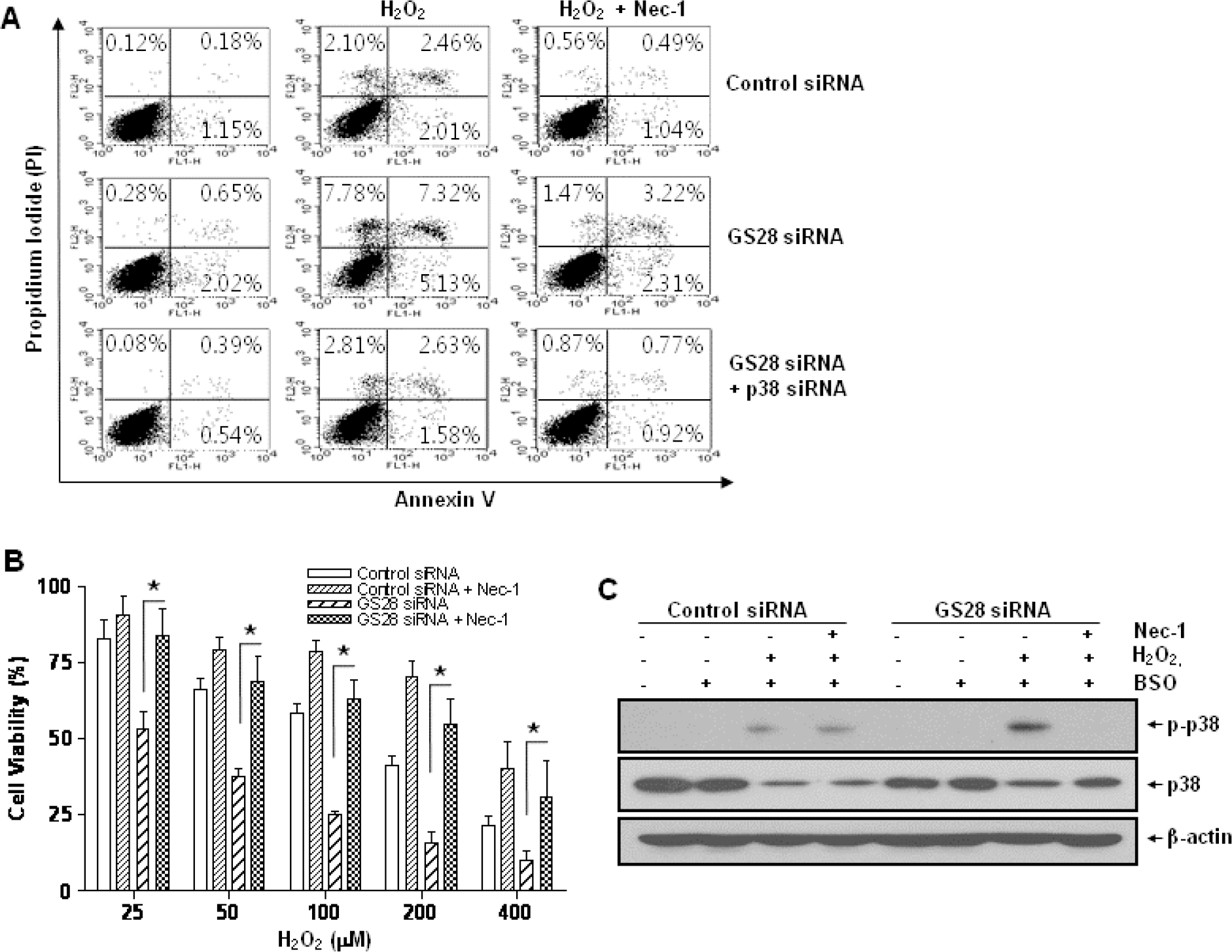 | Fig. 7.H2O2-induced necroptotic cell death via activations of p38 MAPK in GS28 siRNA-transfected cells. (A) Pattern of H2O2-induced cell death was examined in GS28 siRNA-transfected cells without or with cointroduction of p38 siRNA. siRNA-transfected cells were treated with H2O2 for 3 h in the presence of BSO without or with pretreatment with 40 μM necrostatin-1 (Nec-1). The cells were stained with annexin V and propidium iodide (PI) and analyzed by flow cytometry, as described in Materials and Methods. (B) H2O2-induced cell death was examined in GS28 siRNA-transfected cells without or with pretreatment with 40 μM Nec-1. The cells were treated with H2O2 for 24 h in the presence of BSO. Cell viability was determined by an MTT reduction assay. The data represent the mean±SEM of 3 independent experiments. ∗p<0.05 versus the values in the cells transfected with GS28 siRNA alone. (C) H2O2-induced necroptotosis via activations of p38 MAPK was examined in GS28 siRNA-transfected cells. siRNA-transfected cells were treated with 100 μM H2O2 for 3 h without or with pretreatment with 40 μM Nec-1. Phosphorylation of p38 was determined by immunoblot analysis. β-actin was used as a loading control. |
 XML Download
XML Download