

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Abstract
Phenolic compounds affect intracellular free Ca2+ concentration ([Ca2+]i) signaling. The study examined whether the simple phenolic compound octyl gallate affects ATP-induced Ca2+ signaling in PC12 cells using fura-2-based digital Ca2+ imaging and whole-cell patch clamping. Treatment with ATP (100 μM) for 90 s induced increases in [Ca2+]i in PC12 cells. Pretreatment with octyl gallate (100 nM to 20 μM) for 10 min inhibited the ATP-induced [Ca2+]i response in a concentration-dependent manner (IC50=2.84 μM). Treatment with octyl gallate (3 μM) for 10 min significantly inhibited the ATP-induced response following the removal of extracellular Ca2+ with nominally Ca2+-free HEPES HBSS or depletion of intracellular Ca2+ stores with thapsigargin (1 μM). Treatment for 10 min with the L-type Ca2+ channel antagonist nimodipine (1 μM) significantly inhibited the ATP-induced [Ca2+]i increase, and treatment with octyl gallate further inhibited the ATP-induced response. Treatment with octyl gallate significantly inhibited the [Ca2+]i increase induced by 50 mM KCl. Pretreatment with protein kinase C inhibitors staurosporin (100 nM) and GF109203X (300 nM), or the tyrosine kinase inhibitor genistein (50 μM) did not significantly affect the inhibitory effects of octyl gallate on the ATP-induced response. Treatment with octyl gallate markedly inhibited the ATP-induced currents. Therefore, we conclude that octyl gallate inhibits ATP-induced [Ca2+]i increase in PC12 cells by inhibiting both non-selective P2X receptor-mediated influx of Ca2+ from extracellular space and P2Y receptor-induced release of Ca2+ from intracellular stores in protein kinase-independent manner. In addition, octyl gallate inhibits the ATP-induced Ca2+ responses by inhibiting the secondary activation of voltage-gated Ca2+ channels.
Go to : 
References
1. Jahr CE, Jessell TM. ATP excites a subpopulation of rat dorsal horn neurones. Nature. 1983; 304:730–733.

2. Inoue K. ATP receptors for the protection of hippocampal functions. Jpn J Pharmacol. 1998; 78:405–410.
3. Fasolato C, Pizzo P, Pozzan T. Receptor-mediated calcium influx in PC12 cells. ATP and bradykinin activate two independent pathways. J Biol Chem. 1990; 265:20351–20355.
4. Murrin RJ, Boarder MR. Neuronal “nucleotide” receptor linked to phospholipase C and phospholipase D: Stimulation of PC12 cells by ATP analogues and UTP. Mol Pharmacol. 1992; 41:561–568.
5. Kim HJ, Choi JS, Lee YM, Shim EY, Hong SH, Kim MJ, Min DS, Rhie DJ, Kim MS, Jo YH, Hahn SJ, Yoon SH. Fluoxetine inhibits ATP-induced [Ca2+]i increase in PC12 cells by inhibiting both extracellular Ca2+ influx and Ca2+ release from intracellular stores. Neuropharmacology. 2005; 49:265–274.
6. Manach C, Mazur A, Scalbert A. Polyphenols and prevention of cardiovascular diseases. Curr Opin Lipidol. 2005; 16:77–84.
7. Duthie SJ. Berry phytochemicals, genomic stability and cancer: evidence for chemoprotection at several stages in the carcinogenic process. Mol Nutr Food Res. 2007; 51:665–674.
8. Levites Y, Amit T, Mandel S, Youdim MB. Neuroprotection and neurorescue against Abeta toxicity and PKC-dependent release of nonamyloidogenic soluble precursor protein by green tea polyphenol (–)-epigallocatechin-3-gallate. FASEB J. 2003; 17:952–954.
9. Ono K, Yoshiike Y, Takashima A, Hasegawa K, Naiki H, Yamada M. Potent anti-amyloidogenic and fibril-destabilizing effects of polyphenols in vitro: implications for the prevention and therapeutics of Alzheimer's disease. J Neurochem. 2003; 87:172–181.
10. Ramassamy C. Emerging role of polyphenolic compounds in the treatment of neurodegenerative diseases: a review of their intracellular targets. Eur J Pharmacol. 2006; 545:51–64.
11. Silva JP, Proenca F, Coutinho OP. Protective role of new nitrogen compounds on ROS/RNS-mediated damage to PC12 cells. Free Radic Res. 2008; 42:57–69.
12. Zhao B. Natural antioxidants protect neurons in Alzheimer's disease and Parkinson's disease. Neurochem Res. 2009; 34:630–638.
13. Bastianetto S, Yao ZX, Papadopoulos V, Quirion R. Neuro-protective effects of green and black teas and their catechin gallate esters against beta-amyloid-induced toxicity. Eur J Neurosci. 2006; 23:55–64.
14. Ko FN, Huang TF, Teng CM. Vasodilatory action mechanisms of apigenin isolated from Apium graveolens in rat thoracic aorta. Biochim Biophys Acta. 1991; 1115:69–74.
15. Saponara S, Sgaragli G, Fusi F. Quercetin as a novel activator of L-type Ca2+ channels in rat tail artery smooth muscle cells. Br J Pharmacol. 2002; 135:1819–1827.
16. Losi G, Puia G, Garzon G, de Vuono MC, Baraldi M. Apigenin modulates GABAergic and glutamatergic transmission in cultured cortical neurons. Eur J Pharmacol. 2004; 502:41–46.
17. Kim HJ, Yum KS, Sung JH, Rhie DJ, Kim MJ, Min do S, Hahn SJ, Kim MS, Jo YH, Yoon SH. Epigallocatechin-3-gallate increases intracellular [Ca2+]i in U87 cells mainly by influx of extracellular Ca2+ and partly by release of intracellular stores. Naunyn Schmiedebergs Arch Pharmacol. 2004; 369:260–267.
18. Formica JV, Regelson W. Review of the biology of quercetin and related bioflavonoids. Food Chem Toxicol. 1995; 33:1061–1080.
19. Han JH, Kim KJ, Jang HJ, Jang JH, Kim MJ, Sung KW, Rhie DJ, Jo YH, Hahn SJ, Lee MY, Yoon SH. Effects of apigenin on glutamate-induced [Ca2+]i Increases in aultured rat hippocampal neurons. Kor J Physiol Pharmacol. 2008; 12:43–50.
20. Lin F, Xin Y, Wang J, Ma L, Liu J, Liu C, Long L, Wang F, Jin Y, Zhou J, Chen J. Puerarin facilitates Ca2+-induced Ca2+ release triggered by KCl-depolarization in primary cultured rat hippocampal neurons. Eur J Pharmacol. 2007; 570:43–49.
21. Ban JY, Jeon SY, Bae K, Song KS, Seong YH. Catechin and epicatechin from Smilacis chinae rhizome protect cultured rat cortical neurons against amyloid beta protein (25–35)-induced neurotoxicity through inhibition of cytosolic calcium elevation. Life Sci. 2006; 79:2251–2259.
22. Shimmyo Y, Kihara T, Akaike A, Niidome T, Sugimoto H. Three distinct neuroprotective functions of myricetin against glutamate-induced neuronal cell death: involvement of direct inhibition of caspase-3. J Neurosci Res. 2008; 86:1836–1845.
23. Summanen J, Vuorela P, Rauha JP, Tammela P, Marjamaki K, Pasternack M, Tornquist K, Vuorela H. Effects of simple aromatic compounds and flavonoids on Ca2+ fluxes in rat pituitary GH4C1 cells. Eur J Pharmacol. 2001; 414:125–133.
24. Majid MA, Okajima F, Kondo Y. Characterization of ATP receptor which mediates norepinephrine release in PC12 cells. Biochim Biophys Acta. 1992; 1136:283–289.
25. Nakazawa K, Inoue K. Roles of Ca2+ influx through ATP-activated channels in catecholamine release from pheochromocytoma PC12 cells. J Neurophysiol. 1992; 68:2026–2032.
26. Suh BC, Lee CO, Kim KT. Signal flows from two phospholipase C-linked receptors are independent in PC12 cells. J Neurochem. 1995; 64:1071–1079.
27. Sun AY, Chen YM. Extracellular ATP-induced apoptosis in PC12 cells. Adv Exp Med Biol. 1998; 446:73–83.
28. Rhie DJ, Sung JH, Kim HJ, Ha US, Min DS, Kim MS, Jo YH, Hahn SJ, Yoon SH. Endogenous somatostatin receptors mobilize calcium from inositol 1,4,5-trisphosphate-sensitive stores in NG108–15 cells. Brain Res. 2003; 975:120–128.
29. Choi JS, Choi BH, Ahn HS, Kim MJ, Rhie DJ, Yoon SH, Min do S, Jo YH, Kim MS, Sung KW, Hahn SJ. Mechanism of block by fluoxetine of 5-hydroxytryptamine3 (5-HT3)-mediated currents in NCB-20 neuroblastoma cells. Biochem Pharmacol. 2003; 66:2125–2132.
30. Thastrup O, Cullen PJ, Drobak BK, Hanley MR, Dawson AP. Thapsigargin, a tumor promoter, discharges intracellular Ca2+ stores by specific inhibition of the endoplasmic reticulum Ca2+-ATPase. Proc Natl Acad Sci U S A. 1990; 87:2466–2470.
31. Agullo G, Gamet-Payrastre L, Manenti S, Viala C, Remesy C, Chap H, Payrastre B. Relationship between flavonoid structure and inhibition of phosphatidylinositol 3-kinase: a comparison with tyrosine kinase and protein kinase C inhibition. Biochem Pharmacol. 1997; 53:1649–1657.
32. Munaron L, Distasi C, Carabelli V, Baccino FM, Bonelli G, Lovisolo D. Sustained calcium influx activated by basic fibroblast growth factor in Balb-c 3T3 fibroblasts. J Physiol. 1995; 484:557–566.
33. Nakazawa K, Fujimori K, Takanaka A, Inoue K. An ATP-activated conductance in pheochromocytoma cells and its suppression by extracellular calcium. J Physiol. 1990; 428:257–272.
34. Gollasch M, Haller H. Multiple pathways for ATP-induced intracellular calcium elevation in pheochromocytoma (PC12) cells. Ren Physiol Biochem. 1995; 18:57–65.
35. Ferriola PC, Cody V, Middleton E Jr. Protein kinase C inhibition by plant flavonoids. Kinetic mechanisms and structure-activity relationships. Biochem Pharmacol. 1989; 38:1617–1624.
36. Duarte J, Perez Vizcaino F, Utrilla P, Jimenez J, Tamargo J, Zarzuelo A. Vasodilatory effects of flavonoids in rat aortic smooth muscle. Structure-activity relationships. Gen Pharmacol. 1993; 24:857–862.
37. Revuelta MP, Cantabrana B, Hidalgo A. Depolarization-dependent effect of flavonoids in rat uterine smooth muscle contraction elicited by CaCl2. Gen Pharmacol. 1997; 29:847–857.
38. D'Ambrosi N, Murra B, Cavaliere F, Amadio S, Bernardi G, Burnstock G, Volonte C. Interaction between ATP and nerve growth factor signalling in the survival and neuritic outgrowth from PC12 cells. Neuroscience. 2001; 108:527–534.
39. Avallone R, Zanoli P, Puia G, Kleinschnitz M, Schreier P, Baraldi M. Pharmacological profile of apigenin, a flavonoid isolated from Matricaria chamomilla. Biochem Pharmacol. 2000; 59:1387–1394.
40. Paillart C, Carlier E, Guedin D, Dargent B, Couraud F. Direct block of voltage-sensitive sodium channels by genistein, a tyrosine kinase inhibitor. J Pharmacol Exp Ther. 1997; 280:521–526.
41. Choi BH, Choi JS, Min DS, Yoon SH, Rhie DJ, Jo YH, Kim MS, Hahn SJ. Effects of (–)-epigallocatechin-3-gallate, the main component of green tea, on the cloned rat brain Kv1.5 potassium channels. Biochem Pharmacol. 2001; 62:527–535.
42. Hendrich AB, Malon R, Pola A, Shirataki Y, Motohashi N, Michalak K. Differential interaction of Sophora isoflavonoids with lipid bilayers. Eur J Pharm Sci. 2002; 16:201–208.
43. Hendrich AB. Flavonoid-membrane interactions: possible consequences for biological effects of some polyphenolic compounds. Acta Pharmacol Sin. 2006; 27:27–40.
44. Tillman TS, Cascio M. Effects of membrane lipids on ion channel structure and function. Cell Biochem Biophys. 2003; 38:161–190.
45. Zhang YX, Yamashita H, Ohshita T, Sawamoto N, Nakamura S. ATP increases extracellular dopamine level through stimulation of P2Y purinoceptors in the rat striatum. Brain Res. 1995; 691:205–212.
46. Cunha RA, Ribeiro JA. ATP as a presynaptic modulator. Life Sci. 2000; 68:119–137.
47. Choi YM, Jang JY, Jang M, Kim SH, Kang YK, Cho H, Chung S, Park MK. Modulation of firing activity by ATP in dopamine neurons of the rat substantia nigra pars compacta. Neuroscience. 2009; 160:587–595.
Go to :
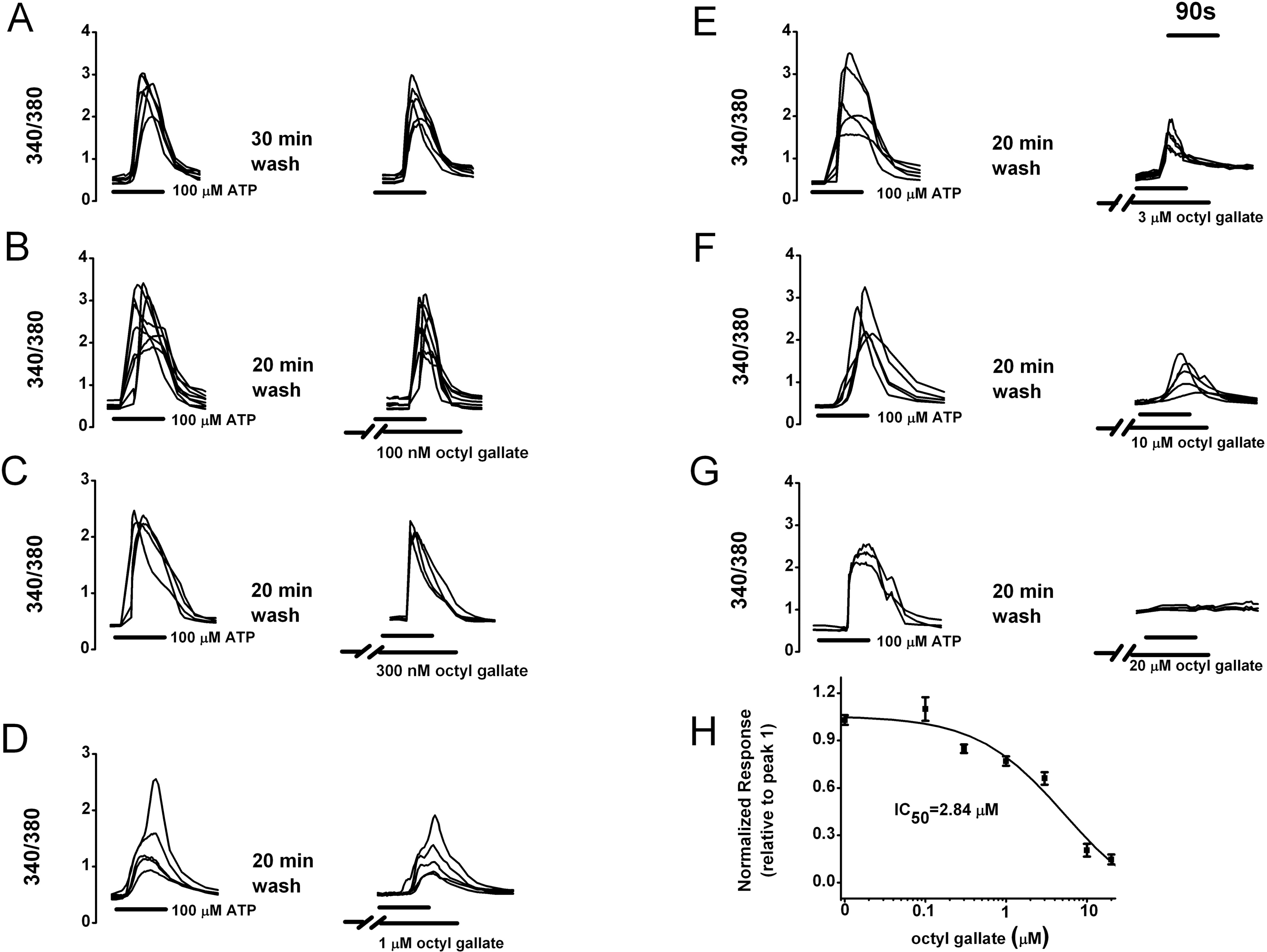 | Fig. 1.Concentration-dependent inhibitory effects of octyl gallate on the ATP-induced [Ca2+]i increase in PC12 cells. (A∼G) After pretreating cells with various concentration of octyl gallate, subsequent ATP-induced [Ca2+]i response was observed. Image pairs were collected at 3∼60 s intervals. ATP and octyl gallate were applied as indicated by the horizontal bars. (H) Summary of concentration-response data. The ATP-induced response amplitude is presented as a percentage of the initial control (relative to peak 1) (n=99, 35, 26, 30, 39, 21, 15 at 0, 100 nM, 300 nM, 1 μM, 3 μM, 10 μM, 20 μM, respectively). A non-linear least-squares fit by the prism software 5.0 to the concentration-response data yielded an IC50 of 2.84 μM for octyl gallate. Data represent mean±SEM. |
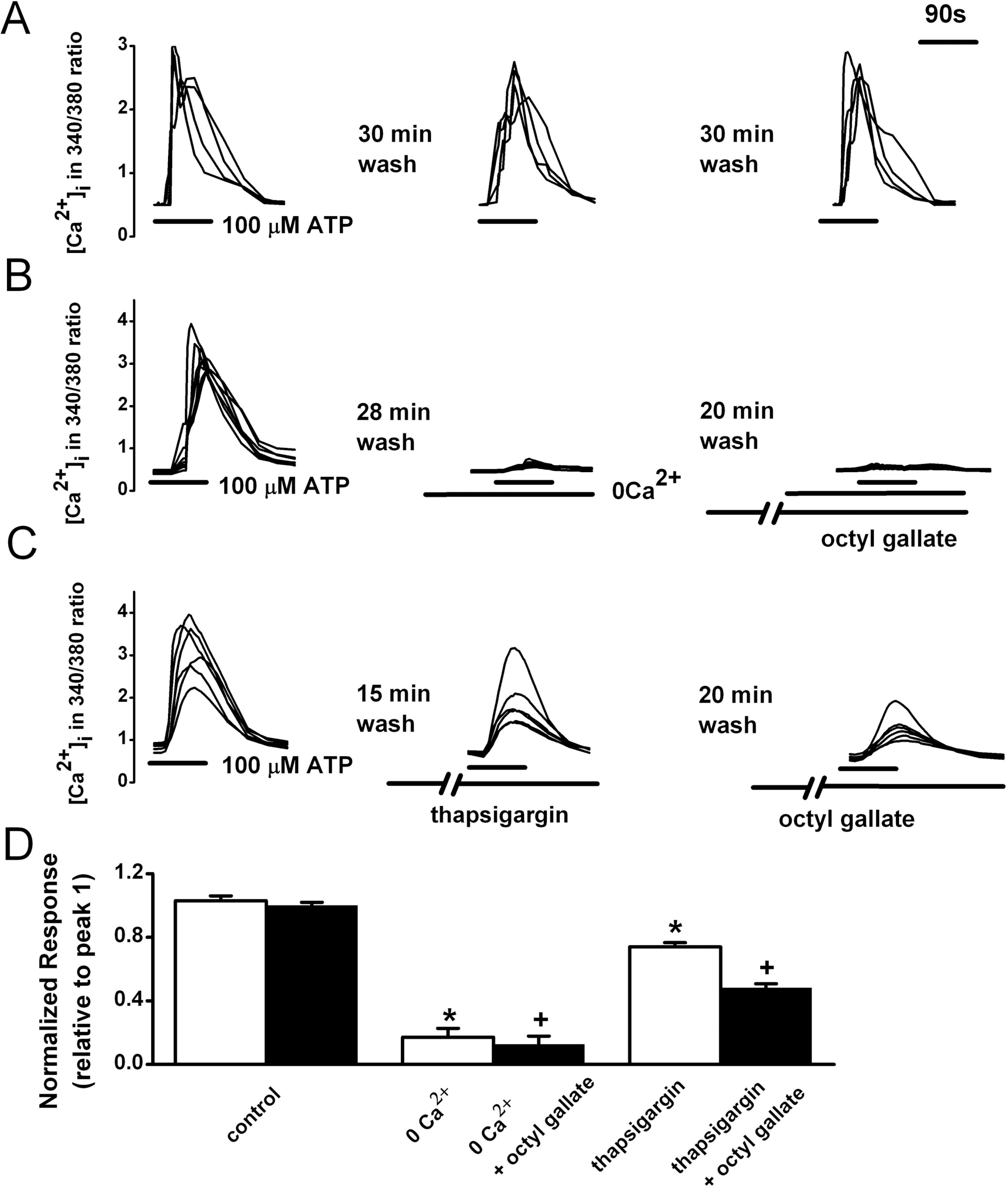 | Fig. 2.Inhibitory effects of octyl gallate on the ATP-induced release of Ca2+ from intracellular stores and Ca2+ influx from the extracellular space. (A) Reproducible [Ca2+]i increases were elicited by super fusion with 100 μM ATP for 90 s at 30 min intervals. ATP-induced [Ca2+]i were recorded after treatment with nominally Ca2+-free HEPES-HBSS for 2 min (B) or with thapsigargin (1 μM) for 15 min (C); then 20 min later the ATP-induced responses were recorded in the presence of octyl gallate (3 μM) for 10 min. (D) Summary of the effects of octyl gallate on ATP-induced [Ca2+]i increase. The amplitude of the ATP-induced response is presented as a percentage of the initial control (relative to peak 1) for the control (n=99), nominally Ca2+-free HEPES-HBSS-treated (0 Ca2+, n=33), 0 Ca2+ plus octyl gallate-treated (n=33), thapsigargin-treated (n=59), and thapsigargin plus octyl gallate-treated (n=59) cells. Data represent mean±SEM. ∗p<0.05 relative to respective control (paired Student's t-test). +p<0.05 relative to respective non-octyl gallate-treated cells (paired Student's t-test). |
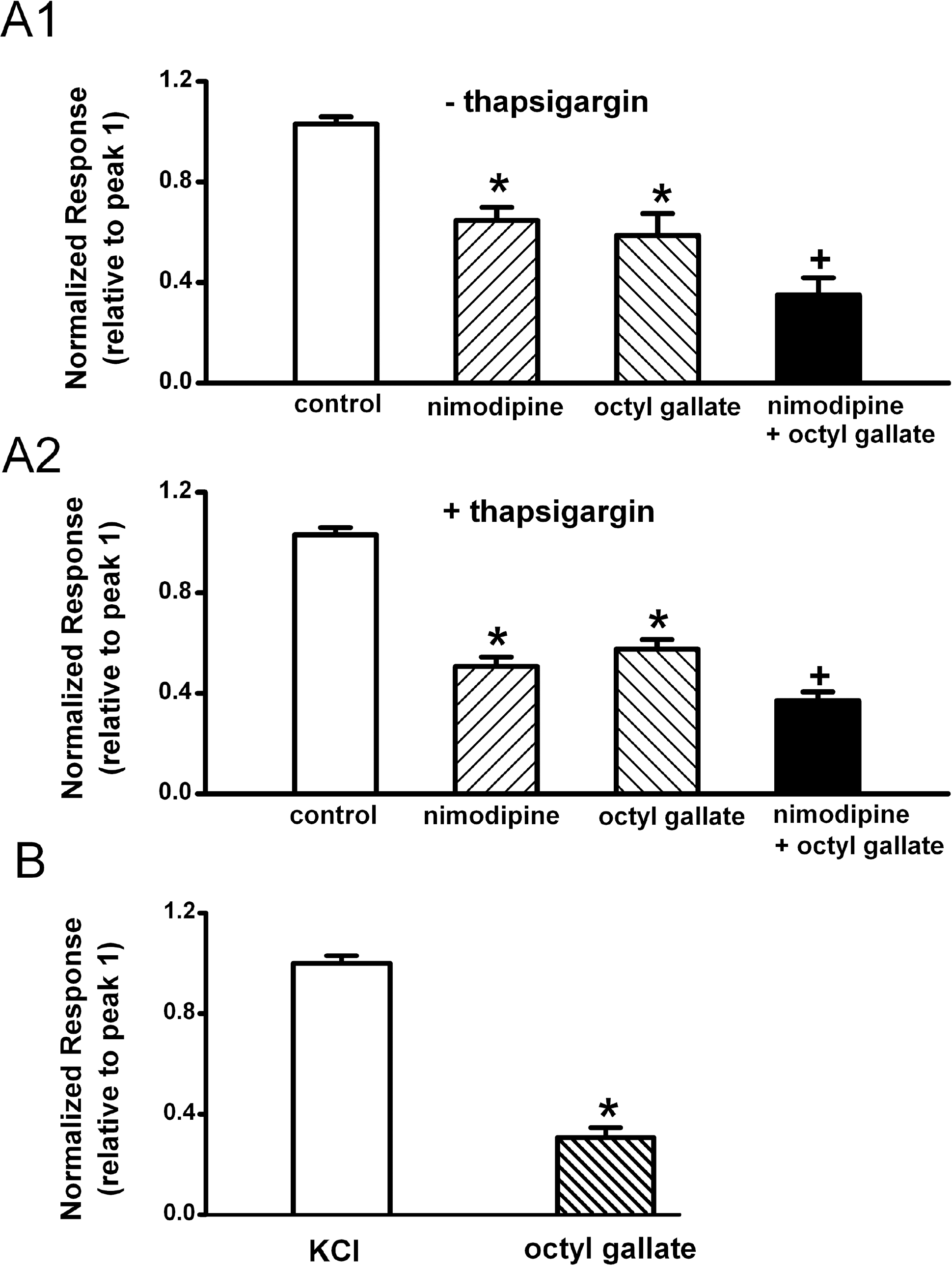 | Fig. 3.Inhibitory effects of octyl gallate on the ATP-induced secondary Ca2+ influx through voltage-gated Ca2+ channels. Effects of nimodipine and octyl gallate on ATP (100 μM)-induced [Ca2+]i increase in untreated cells (A1) and thapsigargin-treated cells (A2). Cells were pretreated with thapsigargin (1 μM) for 45 min during the fura-2 loading period, since thapsigargin-induced increase in [Ca2+]i returns to near basal levels after a 15 min exposure to thapsigargin. The amplitude of the ATP-induced response after treatment of vehicle (control) (n=25, n=24), nimodipine (n=25, n=24), octyl gallate (n=25, n=24), nimodipine plus octyl gallate (n=25, n=24) is presented as a percentage of the initial control in untreated and thapsigargin-treated cells, respectively. (B) Effects of octyl gallate on KCl (50 mM K+ HEPES-HBSS)-induced [Ca2+]i increase. The amplitude of the KCl-induced response after treatment of vehicle (n=31) or octyl gallate (n=26) is presented as a percentage of the initial control. Data represent mean±SEM. ∗p<0.05 relative to respective control or KCl-treated cell; p<0.05 relative to respective nomodipine or octyl gallate-treated cells (one way ANOVA followed by Bonferroni's test and paired Student's t-test). |
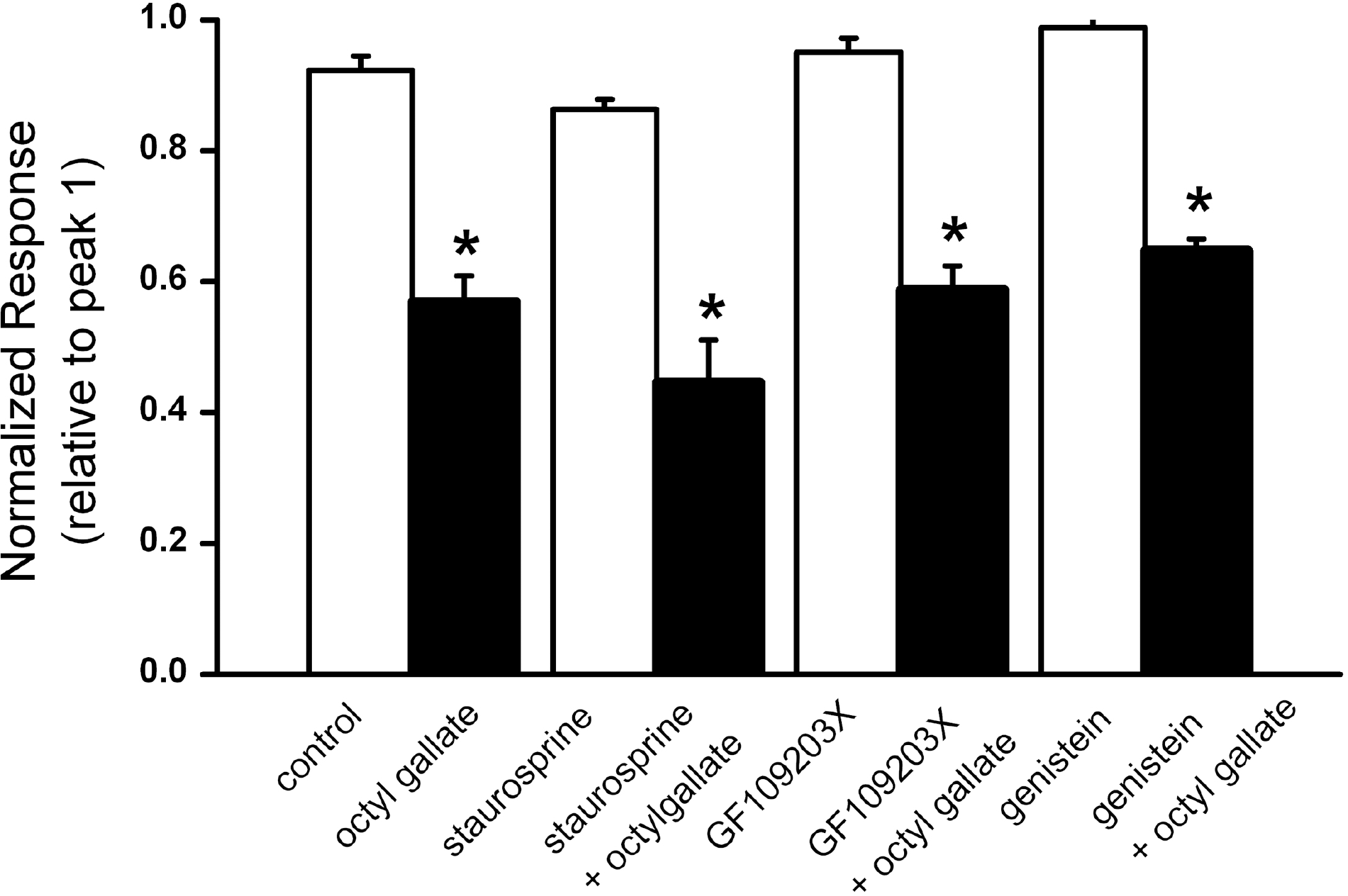 | Fig. 4.ATP-induced [Ca2+]i increase were not inhibited by treatment with PKC inhibitors staurosporin, GF10923X or the tyrosine kinase inhibitor genistein. ATP-induced responses were induced in the presence of vehicle (control, n=59), octyl gallate (3 μM, n=39), staurosprin (100 nM, n=30), staurosprin (100 nM) plus octyl gallate (3 μM) (n=29), GF 109203X (300 nM, n=32), GF 109203X (300 nM) plus octyl gallate (3 μM) (n=29), genistein (50 μM, n=35), genistein (50 μM) plus octyl gallate (3 μM) (n=34) for 10 min following 90 s exposure to ATP and a 20 min wash. The amplitude of the ATP-induced response is presented as a percentage of the initial control responses. Data are expressed as mean±SEM. ∗p<0.05 relative to respective non-octyl gallate-treated cells (one way ANOVA followed by Bonferroni's test). |
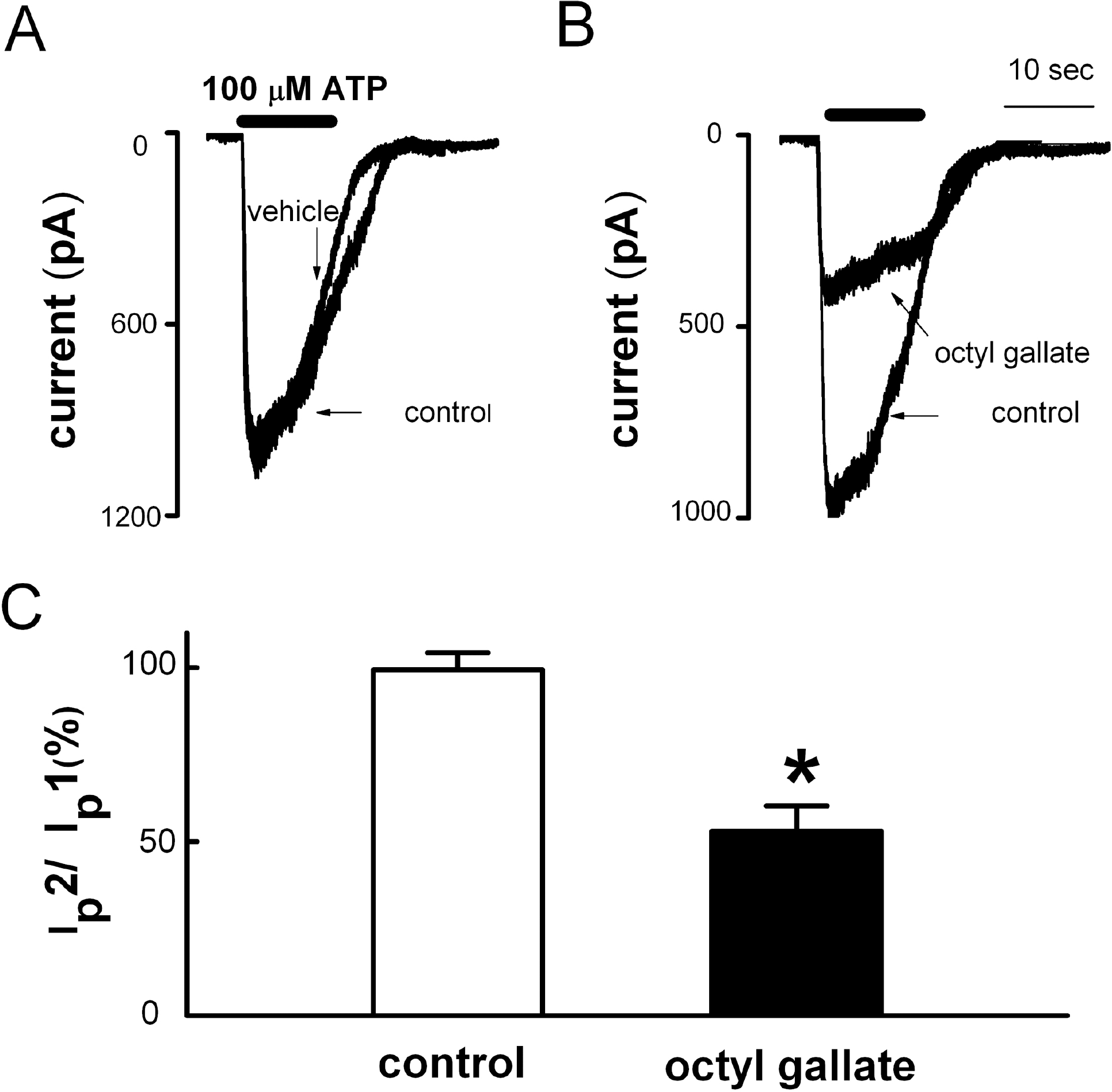 | Fig. 5.Inhibitory effects of octyl gallate on ATP-induced inward currents in PC12 cell. (A) Application of ATP (100 μM, 10 s) evoked inward currents (control). In the same cells, a second application of ATP induced inward current after 10 min washout (vehicle) (n=9). (B) Pretreatment with 3 μM octyl gallate for 10 min inhibited the ATP-induced inward current (octyl gallate, n=6). (C) Summary of the effect of octyl gallate on ATP-induced inward currents. The amplitude of second ATP-induced response (Ip2) is presented as a percentage of the initial control (Ip1) (Ip2/Ip1). Data are expressed as mean±SEM. ∗p<0.05 relative to control (non-paired student's t-test). |
 XML Download
XML Download