

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Abstract
It has been shown that CA repeats in the 3′-untranslated region (UTR) of bcl-2 mRNA contribute the constitutive decay of bcl-2 mRNA and that hnRNP L (heterogenous nuclear ribonucleoprotein L) interacts with CA repeats in the 3′-UTR of bcl-2 mRNA, both in vitro and in vivo. The aim of this study was to determine whether the alteration of hnRNP L affects the stability of bcl-2 mRNA in vivo. Human breast carcinoma MCF-7 cells were transfected with hnRNP L-specific shRNA or hnRNP L-expressing vector to decrease or increase hnRNP L levels, respectively, followed by an actinomycin D chase. An RT-PCR analysis showed that the rate of degradation of endogenous bcl-2 mRNA was not affected by the decrease or increase in the hnRNP L levels. Furthermore, during apoptosis or autophagy, in which bcl-2 expression has been reported to decrease, no difference in the degradation of bcl-2 mRNA was observed between control and hnRNP L-knock down MCF-7 Cells. On the other hand, the levels of AUF-1 and nucleolin, transacting factors for ARE in the 3'UTR of bcl-2 mRNA, were not significantly affected by the decrease in hnRNP L, suggesting that a disturbance in the quantitative balance between these transacting factors is not likely to interfere with the effect of hnRNP L. Collectively, the findings indicate that the decay of bcl-2 mRNA does not appear to be directly controlled by hnRNP L in vivo.
References
1. Heath-Engel HM, Chang NC, Shore GC. The endoplasmic reticulum in apoptosis and autophagy: role of the BCL-2 protein family. Oncogene. 2008; 27:6419–6433.

2. Reed JC. Bcl-2 and the regulation of programmed cell death. J Cell Biol. 1994; 124:1–6.
3. Davies AM. The Bcl-2 family of proteins, and the regulation of neuronal survival. Trends Neurosci. 1995; 18:355–358.
4. Greenlund LJ, Korsmeyer SJ, Johnson EM Jr. Role of BCL-2 in the survival and function of developing and mature sympathetic neurons. Neuron. 1995; 15:649–661.
5. Young RL, Korsmeyer SJ. A negative regulatory element in the bcl-2 5′-untranslated region inhibits expression from an upstream promoter. Mol Cell Biol. 1993; 13:3686–3697.
6. Perillo B, Sasso A, Abbondanza C, Palumbo G. 17beta-estradiol inhibits apoptosis in MCF-7 cells, inducing bcl-2 expression via two estrogen-responsive elements present in the coding sequence. Mol Cell Biol. 2000; 20:2890–2901.
7. Riordan FA, Foroni L, Hoffbrand AV, Mehta AB, Wickremasinghe RG. Okadaic acid-induced apoptosis of HL60 leukemia cells is preceded by destabilization of bcl-2 mRNA and down-regulation of bcl-2 protein. FEBS Lett. 1998; 435:195–198.
8. Bandyopadhyay S, Sengupta TK, Fernandes DJ, Spicer EK. Taxol- and okadaic acid-induced destabilization of bcl-2 mRNA is associated with decreased binding of proteins to a bcl-2 instability element. Biochem Pharmacol. 2003; 66:1151–1162.
9. Guhaniyogi J, Brewer G. Regulation of mRNA stability in mammalian cells. Gene. 2001; 265:11–23.
10. Schiavone N, Rosini P, Quattrone A, Donnini M, Lapucci A, Citti L, Bevilacqua A, Nicolin A, Capaccioli S. A conserved AU-rich element in the 3′ untranslated region of bcl-2 mRNA is endowed with a destabilizing function that is involved in bcl-2 down-regulation during apoptosis. FASEB J. 2000; 14:174–184.
11. Donnini M, Lapucci A, Papucci L, Witort E, Tempestini A, Brewer G, Bevilacqua A, Nicolin A, Capaccioli S, Schiavone N. Apoptosis is associated with modifications of bcl-2 mRNA AU-binding proteins. Biochem Biophys Res Commun. 2001; 287:1063–1069.
12. Lapucci A, Donnini M, Papucci L, Witort E, Tempestini A, Bevilacqua A, Nicolin A, Brewer G, Schiavone N, Capaccioli S. AUF-1 Is a bcl-2 A + U-rich element-binding protein involved in bcl-2 mRNA destabilization during apoptosis. J Biol Chem. 2002; 277:16139–16146.
13. Sengupta TK, Bandyopadhyay S, Fernandes DJ, Spicer EK. Identification of nucleolin as an AU-rich element binding protein involved in bcl-2 mRNA stabilization. J Biol Chem. 2004; 279:10855–10863.
14. Donnini M, Lapucci A, Papucci L, Witort E, Jacquier A, Brewer G, Nicolin A, Capaccioli S, Schiavone N. Identification of TINO: a new evolutionarily conserved BCL-2 AU-rich element RNA-binding protein. J Biol Chem. 2004; 279:20154–20166.
15. Lee JH, Jeon MH, Seo YJ, Lee YJ, Ko JH, Tsujimoto Y, Lee JH. CA repeats in the 3′-untranslated region of bcl-2 mRNA mediate constitutive decay of bcl-2 mRNA. J Biol Chem. 2004; 279:42758–42764.
16. Lee DH, Lim MH, Youn DY, Jung SE, Ahn YS, Tsujimoto Y, Lee JH. hnRNP L binds to CA repeats in the 3'UTR of bcl-2 mRNA. Biochem Biophys Res Commun. 2009; 382:583–587.
17. Park HG, Yoon JY, Choi M. Heterogeneous nuclear ribonucleo-protein D/AUF1 interacts with heterogeneous nuclear ribonucleoprotein L. J Biosci. 2007; 32:1263–1272.
18. Miyashita T, Krajewski S, Krajewska M, Wang HG, Lin HK, Liebermann DA, Hoffman B, Reed JC. Tumor suppressor p53 is a regulator of bcl-2 and bax gene expression in vitro and in vivo. Oncogene. 1994; 9:1799–1805.
19. Suzuki A, Matsuzawa A, Iguchi T. Down regulation of Bcl-2 is the first step on Fas-mediated apoptosis of male reproductive tract. Oncogene. 1996; 13:31–37.
20. Pattingre S, Tassa A, Qu X, Garuti R, Liang XH, Mizushima N, Packer M, Schneider MD, Levine B. Bcl-2 antiapoptotic proteins inhibit Beclin 1-dependent autophagy. Cell. 2005; 122:927–939.
21. Hui J, Reither G, Bindereif A. Novel functional role of CA repeats and hnRNP L in RNA stability. RNA. 2003; 9:931–936.
22. Hui J, Hung LH, Heiner M, Schreiner S, Neumüller N, Reither G, Haas SA, Bindereif A. Intronic CA-repeat and CA-rich elements: a new class of regulators of mammalian alternative splicing. EMBO J. 2005; 24:1988–1998.
23. Rothrock CR, House AE, Lynch KW. HnRNP L represses exon splicing via a regulated exonic splicing silencer. EMBO J. 2005; 24:2792–2802.
24. Shih SC, Claffey KP. Regulation of human vascular endothelial growth factor mRNA stability in hypoxia by heterogeneous nuclear ribonucleoprotein L. J Biol Chem. 1995; 274:1359–1365.
25. Söderberg M, Raffalli-Mathieu F, Lang MA. Identification of a regulatory cis-element within the 3′-untranslated region of the murine inducible nitric oxide synthase (iNOS) mRNA; interaction with heterogeneous nuclear ribonucleoproteins I and L and role in the iNOS gene expression. Mol Immunol. 2007; 44:434–442.
26. Hamilton BJ, Nichols RC, Tsukamoto H, Boado RJ, Pardridge WM, Rigby WF. hnRNP A2 and hnRNP L bind the 3'UTR of glucose transporter 1 mRNA and exist as a complex in vivo. Biochem Biophys Res Commun. 1999; 261:646–651.
Fig. 1.
Down-regulation of hnRNP L expression using shRNA does not affect the degradation of bcl-2 mRNA. (A) MCF-7 cells were transfected with pSUPER.puro vector (vector) or shRNA-hnRNP L (sh-hnRNP L) for 72 h and hnRNP L expression as well as Bcl-2 expression was then examined by western blot. The expression of β-actin was used as a loading control. (B, C) Real-time RT-PCR analysis of hnRNP L mRNA expression or bcl-2 mRNA expression after transfection of sh-hnRNP L for 72 h. The relative levels were normalized to 18s rRNA. The value of vector-transfected MCF-7 cells was arbitrarily designated as 1.0. (D) The decay of endogenous bcl-2 mRNA was measured by Real-time RT-PCR after treatment of actinomycin D. At the indicated time points (0, 2, 4, 6, and 8 h) after addition of actinomycin D (5μg/ml), total RNA was extracted from MCF-7 cells transfected with vector (▪) or with sh-hnRNP L (Δ) and processed for real time PCR as described in Methods. Data were normalized to 18s rRNA and the remaining bcl-2 mRNA is presented as % of initial levels. Values are the mean of three independent experiments±SD.
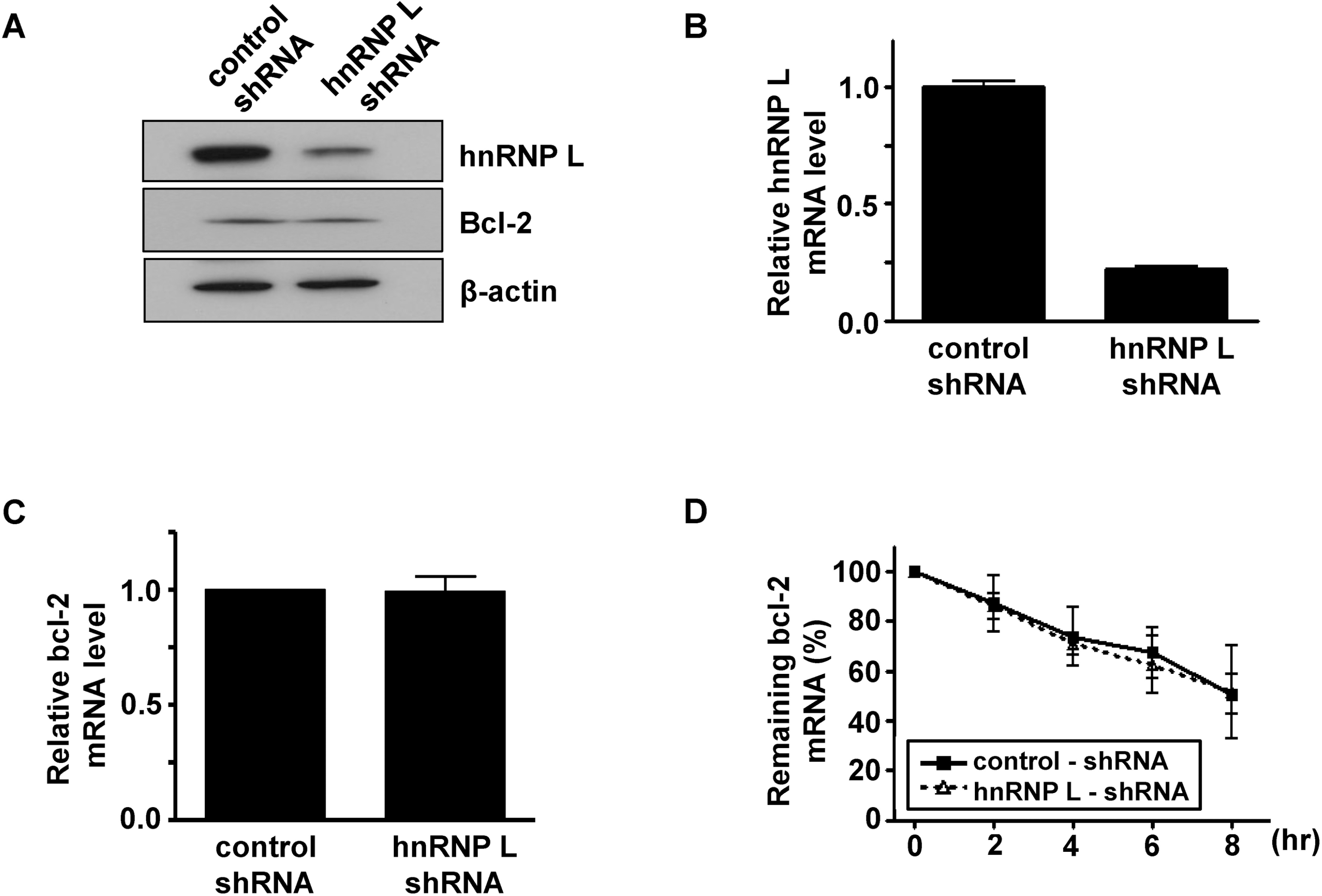
Fig. 2.
Effect of overexpression of hnRNP L on the stability of bcl-2 mRNA in MCF-7 cells. (A) Western blot for hnRNP L expression after transfection of hnRNP L-expressing vector for 48 h. The expression of β-actin was used as a loading control. (B) bcl-2 mRNA half-life was calculated from decay curves by linear regression between 0 and 8 h as in Fig. 1. Values, shown as mean±SD, are based on two independent experiments.
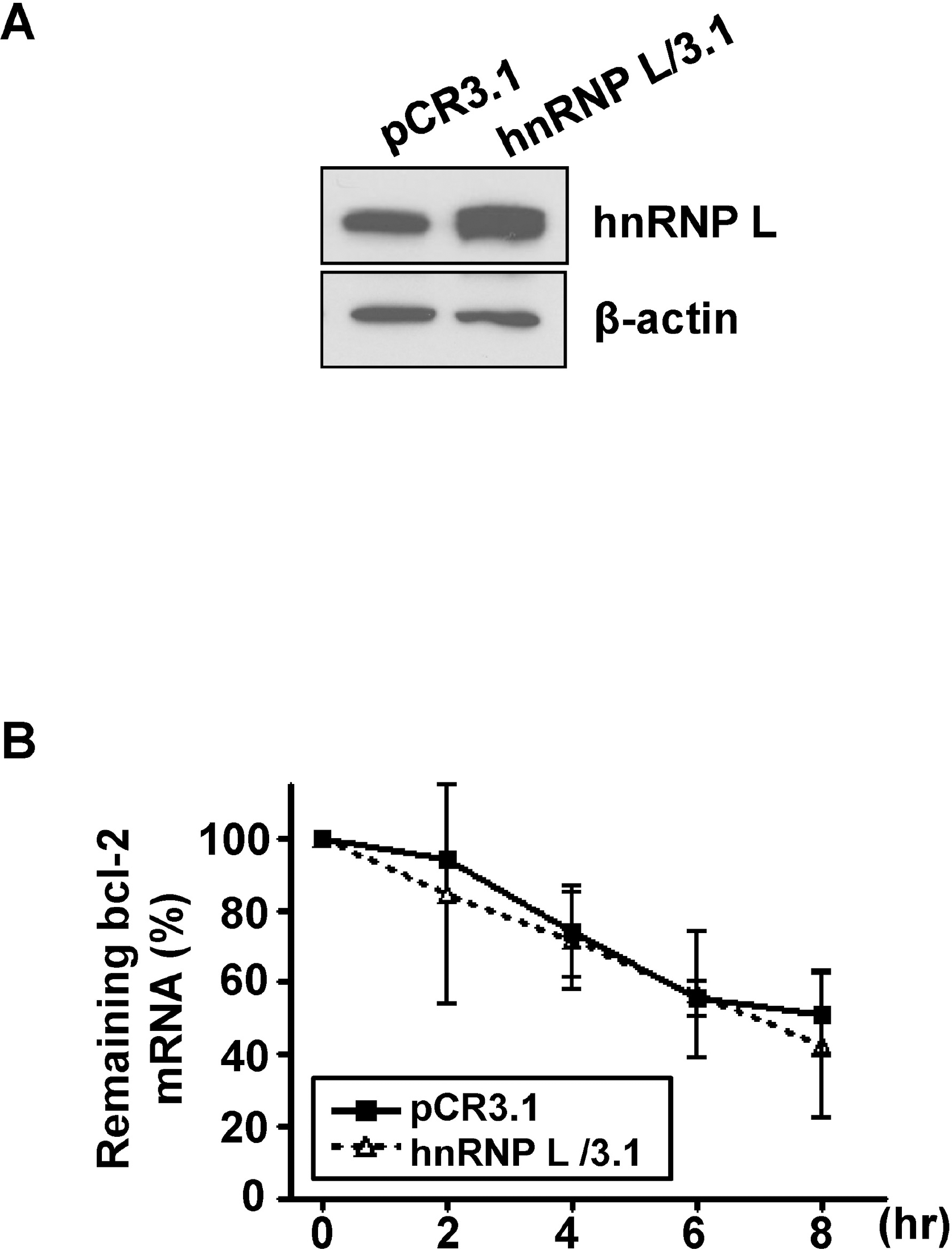
Fig. 3.
Decay of bcl-2 mRNA in vector and sh-hnRNP L-transfected MCF-7 cells during apoptosis or autophagy. After the down-regulation of hnRNP L, MCF-7 cells were exposed to 200 nM of staurospoin (A) or incubated with HBSS (B). Total RNA was isolated at the indicated time points and real-time RT-PCR was performed with bcl-2 and 18s rRNA primers as in Fig. 1. The remaining bcl-2 mRNA at each time point was plotted against time as a percentage of its initial value. Results are shown as means±SD of three independent experiments.
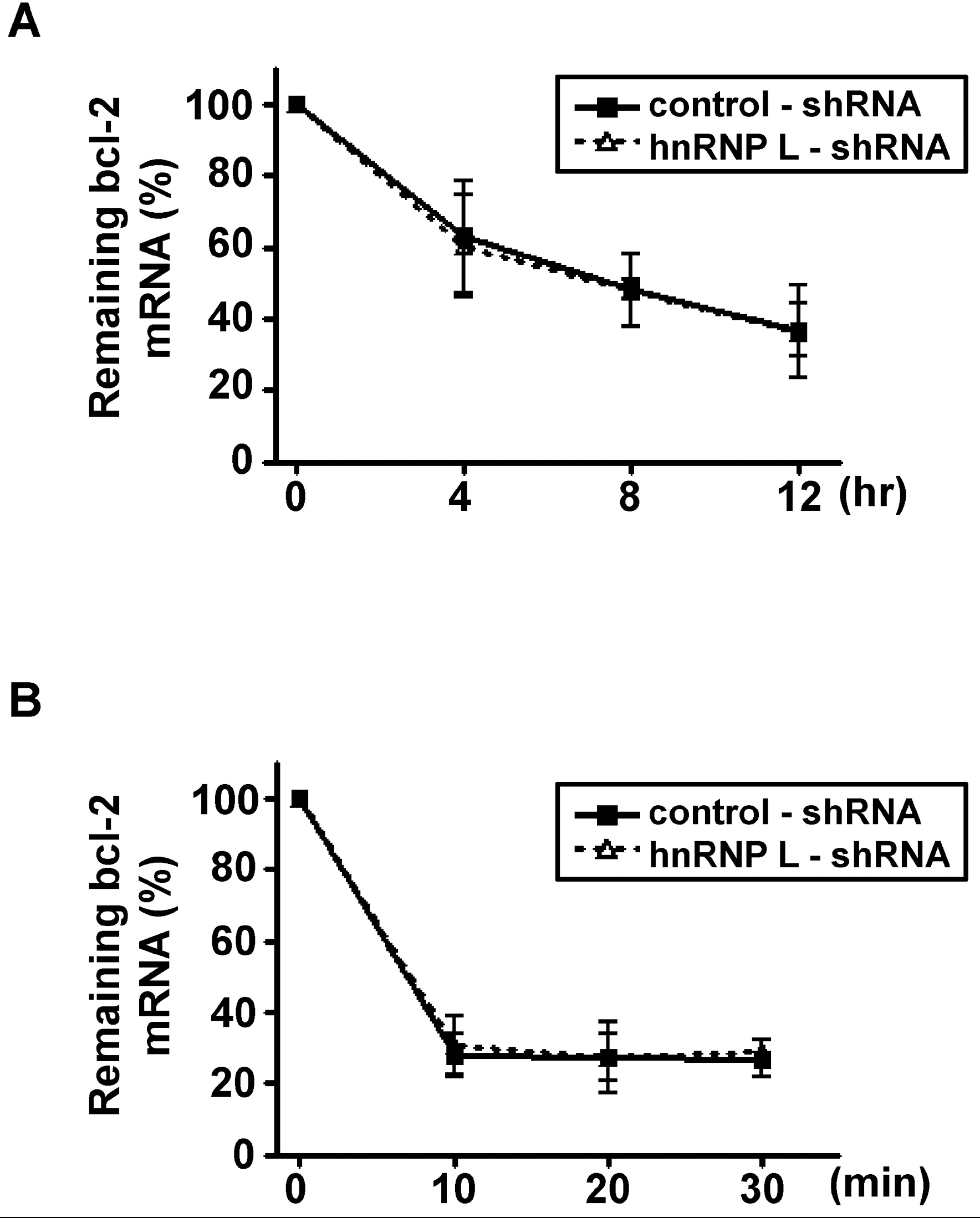
Fig. 4.
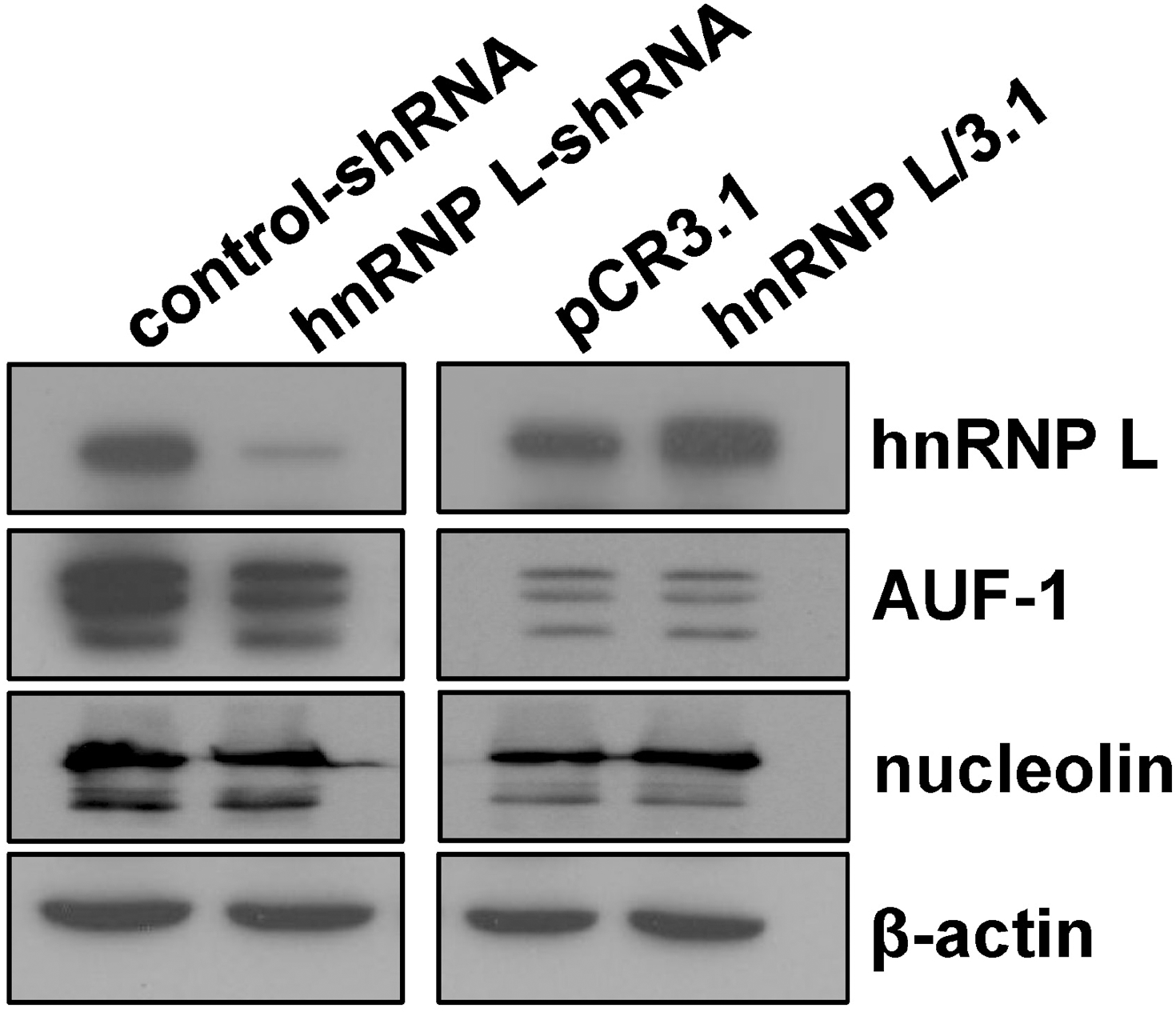
Effect of hnRNP L expression on AUF-1 and nucleolin levels. Total protein lysates were prepared from MCF-7 cells transfected with sh-hnRNP L or hnRNP L-expressing vector. A western blot was performed for hnRNP L, AUF-1 and nucleolin levels. The expression of β-actin was included as a loading control.
 XML Download
XML Download