

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Abstract
Induced activation of the gamma-aminobutyric acidA (GABAA) receptor in the retina of goldfish caused the fish to rotate in the opposite direction to that of the spinning pattern during an optomotor response (OMR) measurement. Muscimol, a GABAA receptor agonist, modified OMR in a concentration-dependent manner. The GABAB receptor agonist baclofen and GABAC receptor agonist CACA did not affect OMR. The observed modifications in OMR included decreased anterograde rotation (0.01∼0.03 μM), coexistence of retrograde rotation and decreased anterograde rotation (0.1∼30 μM) and only retrograde rotation (100 μM∼1 mM). In contrast, the GABAA receptor antagonist bicuculline blocked muscimol-induced retrograde rotation. Based on these results, we inferred that the coding inducing retrograde movement of the goldfish retina is essentially associated with the GABAA receptor-related visual pathway. Furthermore, from our novel approach using observations of goldfish behavior the induced discrete snapshot duration was approximately 573 ms when the fish were under the influence of muscimol.
Go to : 
References
Barlow HB., Hill RM. Selective sensitivity to direction of movement in ganglion cells of the rabbit retina. Science. 139:412–414. 1963.

Barlow HB., Levick WR. The mechanism of directionally selective units in rabbit's retina. J Physiol. 178:477–504. 1965.
Caldwell JH., Daw NW., Wyatt HJ. Effects of picrotoxin and strychnine on rabbit retinal ganglion cells: lateral interactions for cells with more complex receptive fields. J Physiol. 276:277–298. 1978.
Demb JB. Cellular mechanisms for direction selectivity in the retina. Neuron. 55:179–186. 2007.
Egelhaaf M., Borst A. Pilz B. The role of GABA in detecting visual motion. Brain Res. 509:156–160. 1990.
Götz KG. Optomotorische Untersuchung des visuellen Systems einiger Augenmutanten der fruchtfliege Drosophila. Biol Cybern. 2:77–92. 1964.
He S., Masland RH. Retinal direction selectivity after targeted laser ablation of starburst amacrine cells. Nature. 389:378–382. 1997.
Ishida AT., Cohen BN. GABA-activated whole-cell currents in isolated retinal ganglion cells. J Neurophysiol. 60:381–396. 1988.
Kaneko A., Tachibana M. GABA mediates the negative feedback from amacrine to bipolar cells. Neurosci Res Suppl. 6:S239–S251. 1987.
Kittila CA., Massey SC. Effect of ON pathway blockade on directional selectivity in the rabbit retina. J Neurophysiol. 73:703–712. 1995.
Kline K., Holcombe AO., Eagleman DM. Illusory motion reversal is caused by rivalry, not by perceptual snapshots of the visual field. Vision Res. 44:2653–2658. 2004.
Klooster J., Cardozo BN., Yazulla S., Kamermans M. Postsynaptic localization of gamma-aminobutyric acid transporters and receptors in the outer plexiform layer of the goldfish retina: An ultrastructural study. J Comp Neurol. 474:58–74. 2004.
Lin ZS., Yazulla S. Heterogeneity of GABAA receptor in goldfish retina. J Comp Neurol. 345:429–439. 1994.
Maaswinkel H., Li L. Spatio-temporal frequency characteristics of the optomotor response in zebrafish. Vision Res. 43:21–30. 2003.
Mora-Ferrer C., Hausselt S., Hoffmann RS., Ebisch B., Schick S., Wollenberg K., Schneider C., Teege P., Jürgens K. Pharmacological properties of motion vision in goldfish measured with the optomotor response. Brain Res. 1058:17–29. 2005.
Oyster CW., Takahashi E., Collewijn H. Direction-selective retinal ganglion cells and control of optokinetic nystagmus in the rabbit. Vision Res. 12:183–193. 1972.
Paik SS., Park NG., Lee SJ., Han HK., Jung CS., Bai SH., Chun MH. GABA receptors on horizontal cells in the goldfish retina. Vision Res. 43:2101–2106. 2003.
Pakarian P., Yasamy MT. Wagon-wheel illusion under steady illumination: real or illusory? Perception. 32:1307–1310. 2003.
Pan ZH., Slaughter MM. Control of retinal information coding by GABAB receptors. J Neurosci. 11:1810–1821. 1991.
Purves D., Paydarfar JA., Andrews TJ. The wagon wheel illusion in movies and reality. Proc Natl Acad Sci USA. 93:3693–3697. 1996.
Reichardt W. Evaluation of optical motion information by movement detectors. J Comp Physiol A. 161:533–547. 1987.
Schaerer S., Neumeyer C. Motion detection in goldfish investigated with the optomotor response is “color blind”. Vision Res. 36:4025–4034. 1996.
Srinivasan MV., Poteser M., Kral K. Motion detection in insect orientation and navigation. Vision Res. 39:2749–2766. 1999.
VanRullen R. The continuous wagon wheel illusion depends on, but is not identical to neuronal adaptation. Vision Res. 47:2143–2149. 2007.
VanRullen R., Reddy L., Koch C. The continuous wagon wheel illusion is associated with changes in electroencephalogram power at approximately 13 Hz. J Neurosci. 26:502–507. 2006.
Watanabe S., Koizumi A., Matsunaga S., Stocker JW., Kaneko A. GABA-mediated inhibition between amacrine cells in the goldfish retina. J Neurophysiol. 84:1826–1834. 2000.
Werblin FS. Response of retinal cells to moving spots: intracellular recording in Necturus maculosus. J Neurophysiol. 33:342–350. 1969.
Wyatt HJ., Daw NW. Specific effects of neurotransmitter antagonists on ganglion cells in rabbit retina. Science. 191:204–205. 1976.
Yazulla S., Studholme KM., Vitorica J., de Blas AL. Immunocy-tochemical localization of GABAA receptors in goldfish and chicken retinas. J Comp Neurol. 280:15–26. 1989.
Zanker JM., Srinivasan MV., Egelhaaf M. Speed tuning in elementary motion detectors of the correlation type. Biol Cybern. 80:109–116. 1999.
Go to :
 | Fig. 1.OMR of a goldfish before (  ) and after ( ) and after ( ) administration of three concentrations of muscimol (A), baclofen (B), and CACA (C). All drugs were independently administered five times. Muscimol-induced negative OMR represents the reverse rotation of a goldfish to the direction of pattern rotation. Here and in subsequent figures, unless otherwise indicated the error bar represents± standard error of the mean, ∗indicates p<0.05 and n represents the number of goldfish. ) administration of three concentrations of muscimol (A), baclofen (B), and CACA (C). All drugs were independently administered five times. Muscimol-induced negative OMR represents the reverse rotation of a goldfish to the direction of pattern rotation. Here and in subsequent figures, unless otherwise indicated the error bar represents± standard error of the mean, ∗indicates p<0.05 and n represents the number of goldfish. |
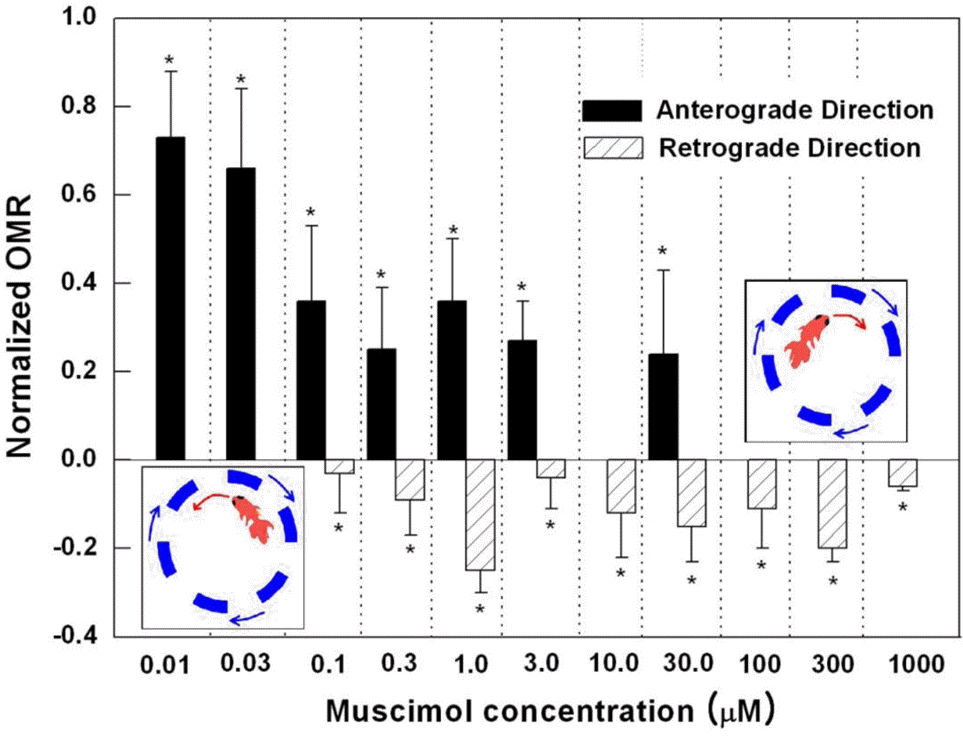 | Fig. 2.The relationship between muscimol concentration and normalized OMR. Eleven concentrations of muscimol from 0.01 μM to 1 mM were used (n=5 for each concentration). At certain concentrations, the same goldfish showed both negative and positive OMR. To account for this, each fish treated with concentrations ranging from 0.1∼30 μM was measured twice as much. Therefore, the total number of fish used in the experiment was 55, but 30 of were tested and measured twice as much as the remaining 25. As the muscimol concentration increased, there were 3 distinctive OMR results: reduction in anterograde rotation, coexistence of retrograde rotation and reduced anterograde rotation, and retrograde rotation only. The diagram, which shows the anterograde and retrograde rotation, is shown on the graph for reference and clarity. |
 | Fig. 3.OMR as to GABA receptor related drugs. (A) Co-injection (n=5) of 1 mM muscimol (MUS) and 2 mM bicuculline (BIC) inhibited the negative OMR that muscimol had induced. (B) OMR using the two concentrations of bicuculline. Bicuculline alone markedly reduced anterograde OMR (n=5). |
 | Fig. 4.Goldfish rotation versus pattern rotation. (A) Number of rotations depending on various pattern velocities after the administration of 1 mM muscimol (n=5 for each velocity). The reversed rotation due to muscimol was the greatest at a pattern velocity of 12 rpm. (B) Rotation of goldfish to low speeds in the control condition (n=5 in different pattern rotation speeds). From this relationship, a linear equation of y=0.565x–0.231 was obtained. Using this, we could predict the pattern's velocity to be 3.12 rpm when a goldfish rotated at a velocity of 1.532 rpm. |
 XML Download
XML Download