

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Abstract
Melatonin has been reported to protect neurons from a variety of neurotoxicity. However, the underlying mechanism by which melatonin exerts its neuroprotective property has not yet been clearly understood. We previously demonstrated that melatonin protected kainic acid-induced neuronal cell death in mouse hippocampus, accompanied by sustained activation of Akt, a critical mediator of neuronal survival. To further elucidate the neuroprotective action of melatonin, we examined in the present study the causal mechanism how Akt signaling pathway is regulated by melatonin in a rat primary astrocyte culture model. Melatonin resulted in increased astrocytic Akt phosphorylation, which was significantly decreased with wortmannin, a specific inhibitor of PI3K, suggesting that activation of Akt by melatonin is mediated through the PI3K-Akt signaling pathway. Furthermore, increased Akt activation was also significantly decreased with luzindole, a non-selective melatonin receptor antagonist. As downstream signaling pathway of Akt activation, increased levels of CREB phoshorylation and GDNF expression were observed, which were also attenuated with wortmannin and luzindole. These results strongly suggest that melatonin exerts its neuroprotective property in astrocytes through the activation of plasma membrane receptors and then PI3K-Akt signaling pathway.
Go to : 
REFERENCES
Anhe GF., Caperuto LC., Pereira-Da-Silva M., Souza LC., Hirata AE., Velloso LA., Cipolla-Neto J., Carvalho CR. In vivo activation of insulin receptor tyrosine kinase by melatonin in the rat hypothalamus. J Neurochem. 90:559–566. 2004.

Baecker PA., Lee WH., Verity AN., Eglen RM., Johnson RM. Characterization of a promoter for the human glial cell line-derived neurotrophic factor gene. Brain Res Mol Brain Res. 69:209–222. 1999.
Boutin JA., Audinot V., Ferry G., Delagrange P. Molecular tools to study melatonin pathways and actions. Trends Pharmacol Sci. 26:412–419. 2005.
Castro LM., Gallant M., Niles LP. Novel targets for valproic acid: up-regulation of melatonin receptors and neurotrophic factors in C6 glioma cells. J Neurochem. 95:1227–1236. 2005.
Conti AG., Maestroni J. The clinical neuroimmunotherapeutic role of melatonin in oncology. J Pineal Res. 19:103–110. 1995.
Du K., Montminy M. CREB is a regulatory target for the protein kinase Akt/PKB. J Biol Chem. 273:32377–32379. 1998.
Henshall DC., Araki T., Schindler CK., Lan JQ., Tiekoter KL. W TakiRP Simon. Activation of Bcl-2-associated death protein and counter-response of Akt within cell populations during seizure-induced neuronal death. J Neurosci. 22:8458–8465. 2002.
Kilic U., Kilic E., Reiter RJ., Bassetti CL., Hermann DM. Signal transduction pathways involved in melatonin-induced neuro-protection after focal cerebral ischemia in mice. J Pineal Res. 38:67–71. 2005.
Kim AH., Yano H., Cho H., Meyer D., Monks B., Margolis B., Birnbaum MJ., Chao MV. Akt1 regulates a JNK scaffold during excitotoxic apoptosis. Neuron. 35:697–709. 2002.
Lee SH., Chun W., Kong PJ., Han JA., Cho BP., Kwon OY., Lee HJ., Kim SS. Sustained activation of Akt by melatonin contributes to the protection against kainic acid-induced neuronal death in hippocampus. J Pineal Res. 40:79–85. 2006.
Lenhard T., Schober A., Suter-Crazzolara C., Unsicker K. Fibroblast growth factor-2 requires glial-cell-line-derived neurotrophic factor for exerting its neuroprotective actions on glutamate- lesioned hippocampal neurons. Mol Cell Neurosci. 20:181–197. 2002.
Maestroni GJ., Conti A., Pierpaoli W. Role of the pineal gland in immunity. Circadian synthesis and release of melatonin modulates the antibody response and antagonizes the immuno-suppressive effect of corticosterone. J Neuroimmunol. 13:19–30. 1986.
Musshoff U., Riewenherm D., Berger E., Fauteck JD., Speckmann EJ. Melatonin receptors in rat hippocampus: molecular and functional investigations. Hippocampus. 12:165–173. 2002.
Peters JL., Earnest BJ., Tjalkens RB., Cassone VM., Zoran MJ. Modulation of intercellular calcium signaling by melatonin in avian and mammalian astrocytes is brain region-specific. J Comp Neurol. 493:370–380. 2005.
Reppert SM., Weaver DR., Godson C. Melatonin receptors step into the light: cloning and classification of subtypes. Trends Pharmacol Sci. 17:100–102. 1996.
Romashkova JA., Makarov SS. NF-kappaB is a target of AKT in anti-apoptotic PDGF signalling. Nature. 401:86–90. 1999.
Tang YP., Ma YL., Chao CC., Chen KY., Lee EH. Enhanced glial cell line-derived neurotrophic factor mRNA expression upon (–)-deprenyl and melatonin treatments. J Neurosci Res. 53:593–604. 1998.
Woodbury D., Schaar DG., Ramakrishnan L., Black IB. Novel structure of the human GDNF gene. Brain Res. 803:95–104. 1998.
Xu Q., Li Y., Cyras C., Sanan DA., Cordell B. Isolation and characterization of apolipoproteins from murine microglia. Identification of a low density lipoprotein-like apolipoprotein J-rich but E-poor spherical particle. J Biol Chem. 275:31770–31777. 2000.
Yano S., Tokumitsu H., Soderling TR. Calcium promotes cell survival through CaM-K kinase activation of the protein-kinase-B pathway. Nature. 396:584–587. 1998.
Young D., Lawlor PA., Leone P., Dragunow M., During MJ. Environmental enrichment inhibits spontaneous apoptosis, prevents seizures and is neuroprotective. Nat Med. 5:448–453. 1999.
Go to :
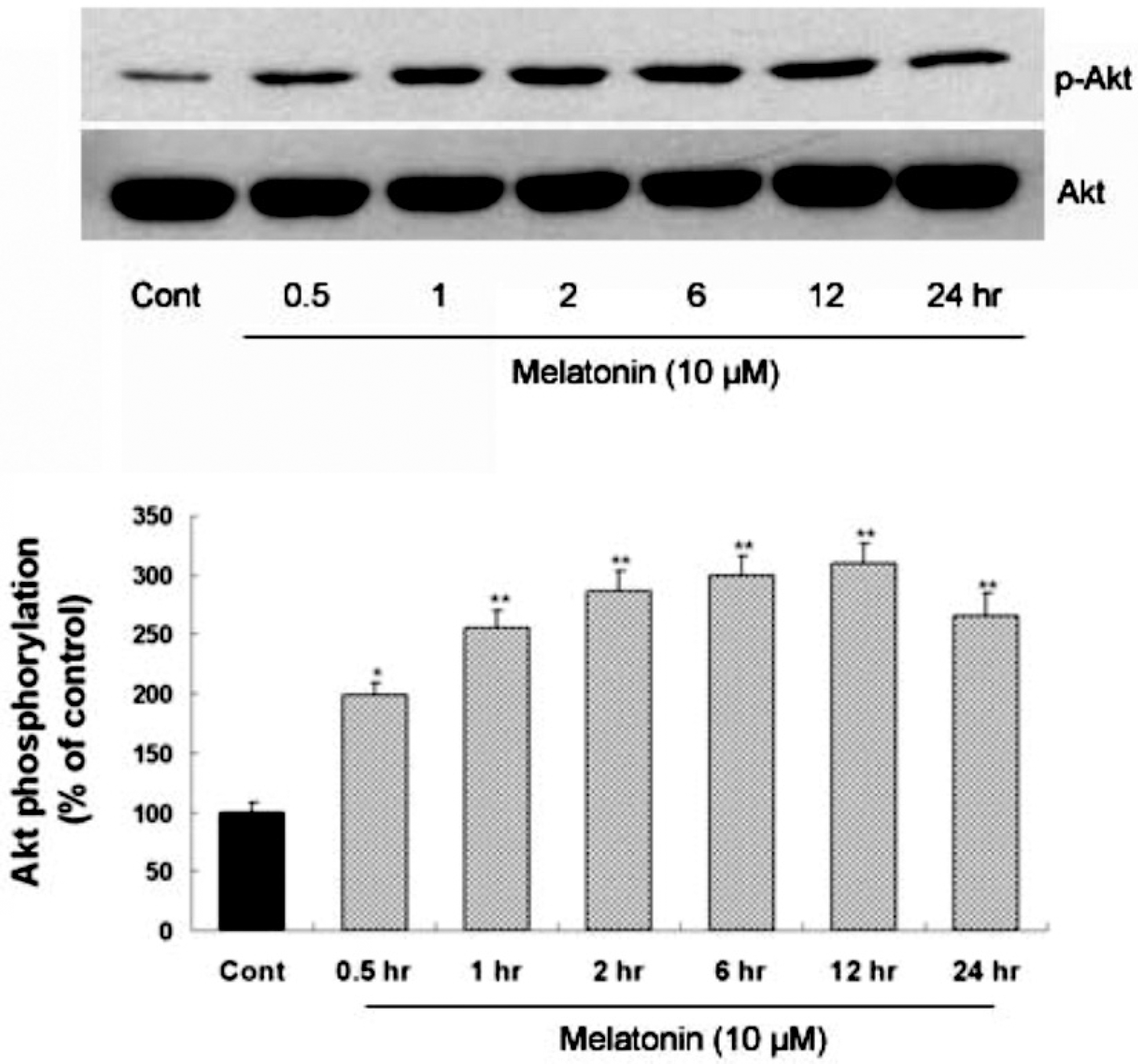 | Fig. 1.Time-course of melatonin-stimulated Akt phosphorylation at Ser473 in primary cultures of rat cortical astrocytes. Quiescent astrocytes were treated with melatonin (10μM) for indicated time periods. Data are representative of four independent experiments. The phospho-Akt and total Akt bands of the samples were densitometrically evaluated and phospho-Akt/total Akt ratio was expressed as mean±SEM. ∗p<0.05 and ∗∗p<0.01 indicate statistically significant difference from the control group. |
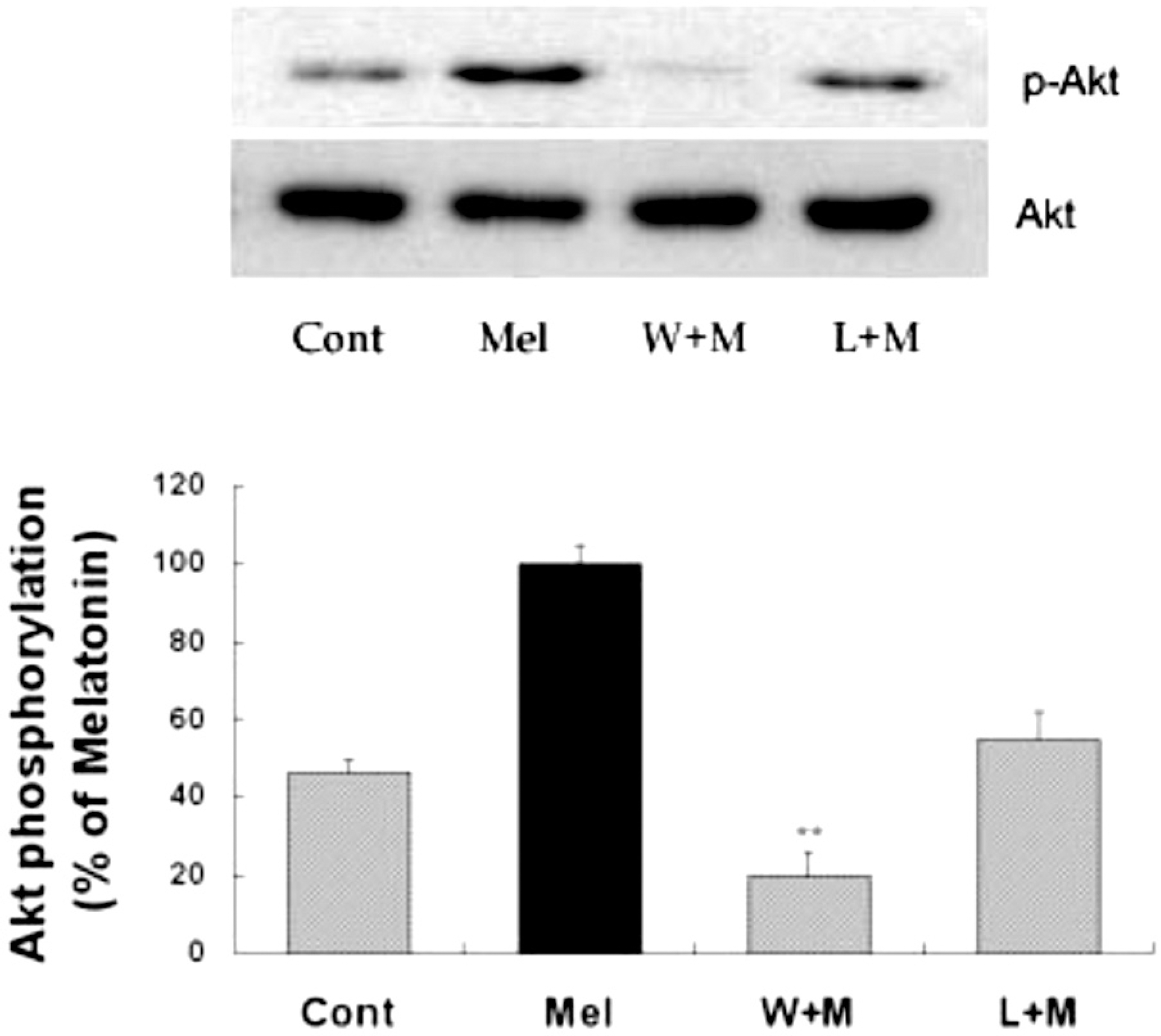 | Fig. 2.Effects of PI3K inhibition and melatonin receptor blockade on melatonin-induced Akt phosphorylation. Melatonin (10μM) was applied to primary cultures for 12 hr in the absence or presence of wortmannin or luzindole. Concentrations and preincubation times of these compounds were as follows: wortmannin (W), 10μM, 1 hr; luzindole (L), 100μM, 1 hr. Right panel represents quantitative analysis. The phospho-Akt/Akt ratio was determined and expressed as mean±SEM. Akt phosphorylation by melatonin was inhibited by pretreatment of the cells with wortmannin or luzindole. ∗p<0.05 and ∗∗p<0.01 indicate statistically significant difference from the melatonin treated group. |
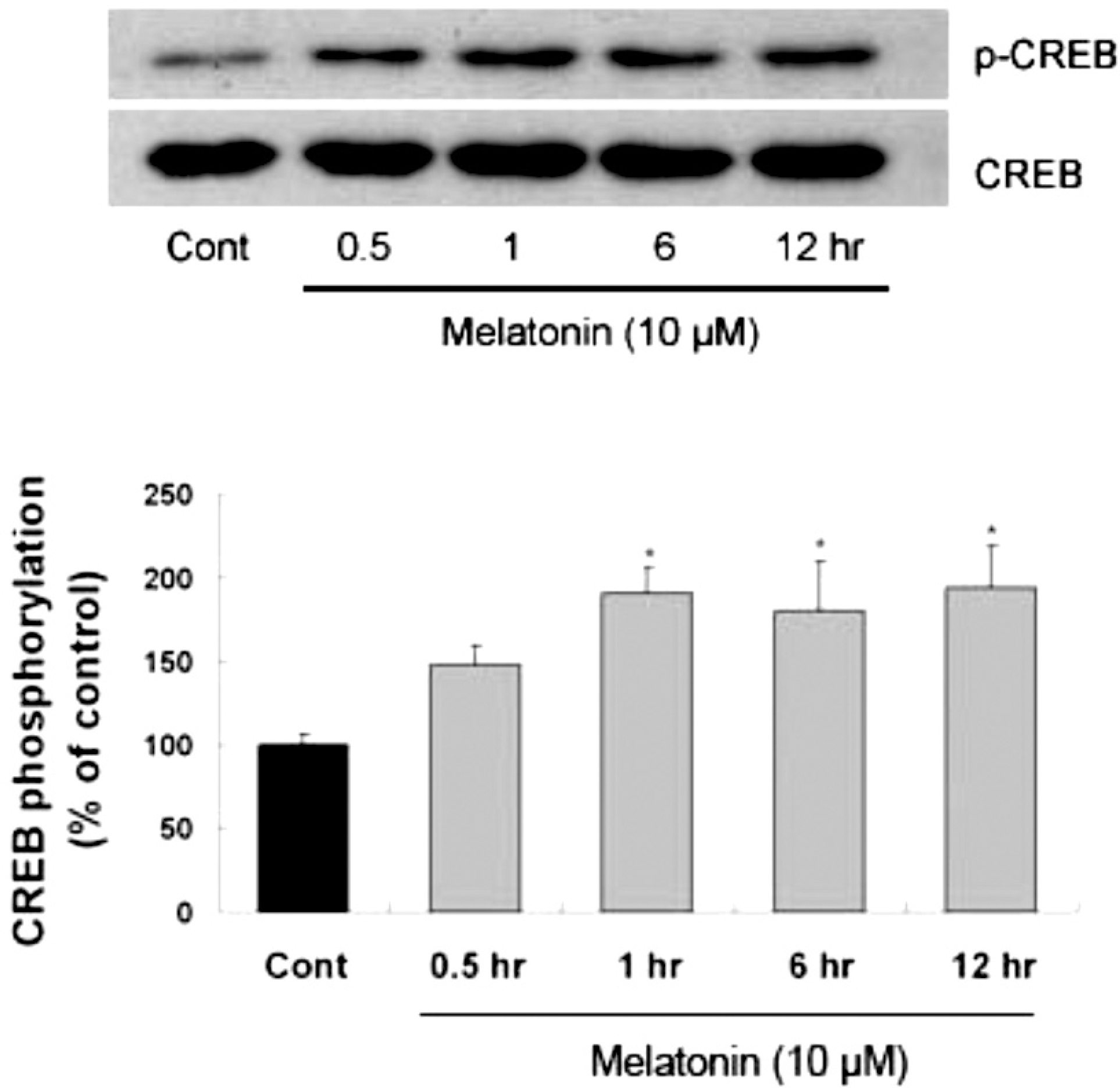 | Fig. 3.Time-course of melatonin-stimulated CREB phosphorylation in primary cultures of rat cortical astrocytes. Quiescent cultured astrocytes were treated with melatonin (10μM) for indicated time periods. CREB phosphorylation in mouse hippocampus was increased by melatonin treatment. Left panel is the representative blot, and right panel shows quantitative verification. The phospho-CREB/CREB ratio was expressed as mean±SEM. ∗p<0.05 indicates statistically significant difference from the control group. |
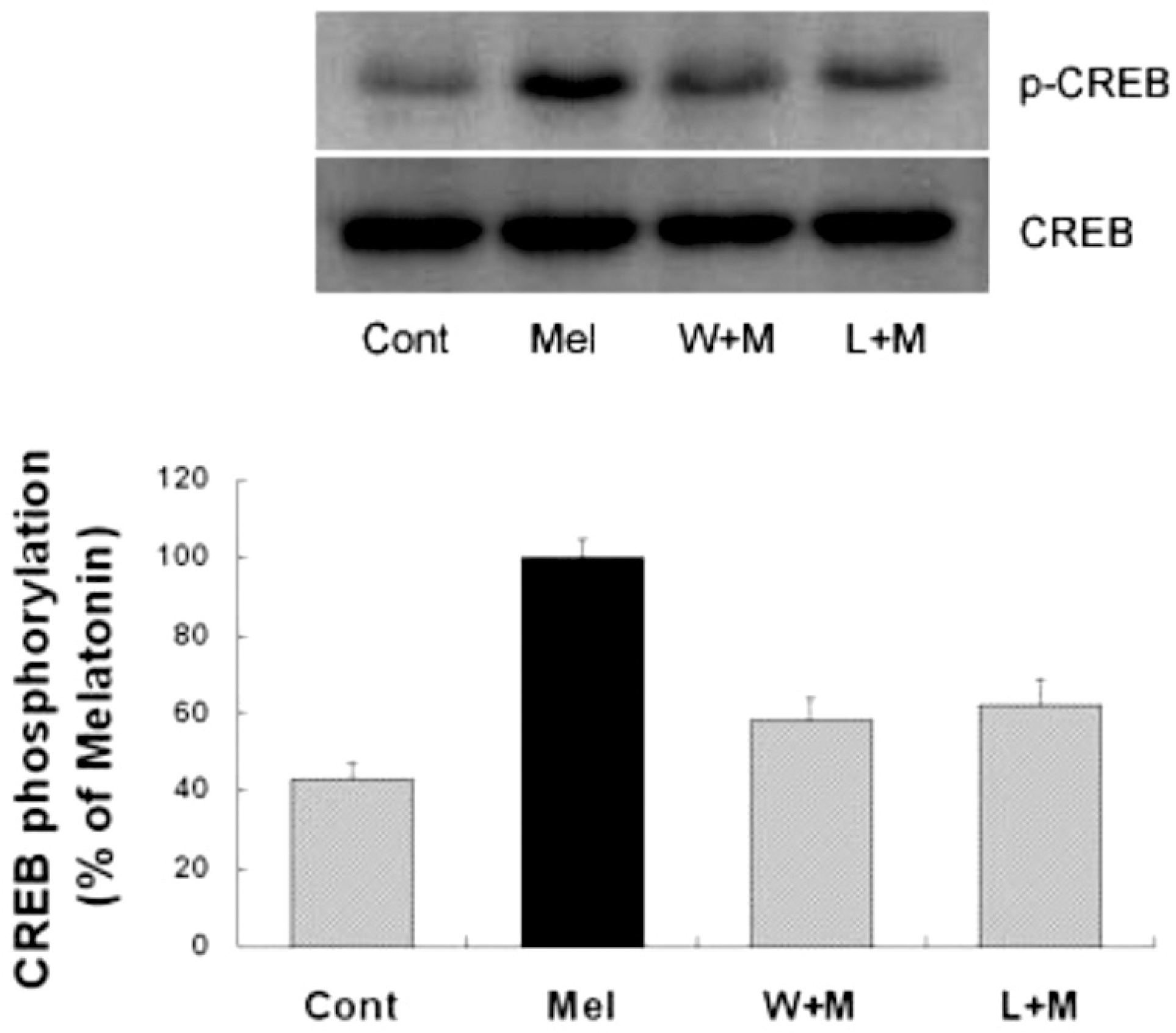 | Fig. 4.Effects of PI3K inhibition and melatonin receptor blockade on melatonin-induced CREB phosphorylation. Melatonin (10μM) was applied to primary cultures for 12 hr in the absence or presence of the inhibitor indicated. Left panel is the representative blot, and right panel shows quantitative verification. The phospho-CREB/CREB ratio was expressed as mean±SEM. CREB phosphorylation by melatonin was partially inhibited by pretreatment of the cells with wortmannin or luzindole. ∗p<0.05 indicates statistically significant difference from the melatonin treated group. |
 | Fig. 5.Effects of PI3K inhibition or melatonin receptor blockade on melatonin-induced neurotrophic factors expression. Melatonin (10 μM) was applied to primary cultures for 24 hr in the absence or presence of the agents indicated. Left panel is a representative blot, and right panel shows quantitative verification. The GDNF/β-actin ratio was expressed as mean±SEM. GDNF expression by melatonin was inhibited by pretreatment of the cells with wortmannin and luzindole. ∗∗p < 0.01 indicates statistically significant difference from the melatonin treated group. |
 XML Download
XML Download