

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Abstract
Caveolin-1 (CAV1) is an integral membrane protein that may function as a scaffold for plasma membrane proteins and acts as a tumor suppressor protein. One causative factor of chemotherapy-resistant cancers is P-plycoprotein (P-gp), the product of the multidrug resistance-1 gene (MDR1), which is localized in the caveolar structure. Currently, the interactive roles of CAV1 and MDR1 expression in the death of cancer cells remain controversial. In this study, we investigated the effects of indomethacin on the cell viability and the expression levels of MDR1 mRNA and protein in a CAV1-siRNA-mediated gene knockdown hepatoma cell line (SK-Hep1). Cell viability was significantly decreased in CAV1-siRNA-transfected cells compared with that of control-siRNA-transfected cells. Furthermore, the viability of cells pretreated with CAV1 siRNA was markedly decreased by treatment with indomethacin (400μM for 24 h). However, the protein and mRNA levels of MDR1 were unchanged in CAV1-siRNA-transfected cells. These results suggest that CAV1 plays an important role as a major survival enzyme in cancer cells, and indomethacin can sensitively induce cell death under conditions of reduced CAV1 expression, independent of MDR1 expression.
Go to : 
REFERENCES
Belanger MM., Gaudreau M., Roussel E., Couet J. Role of caveolin-1 in etoposide resistance development in A549 lung cancer cells. Cancer Biol Ther. 3:954–959. 2004.
Bender FC., Reymond MA., Bron C., Quest AF. Caveolin-1 levels are down-regulated in human colon tumors, and ectopic expression of caveolin-1 in colon carcinoma cell lines reduces cell tumorigenicity. Cancer Res. 60:5870–5878. 2000.
Bruix J., Hessheimer A J., Forner A., Boix L., Vilana R., Llovet JM. New aspects of diagnosis and therapy of hepatocellular carcinoma. Oncogene. 25:3848–3856. 2006.

Cai C., Zhu H., Chen J. Overexpression of caveolin-1 increases plasma membrane fluidity and reduces P-glycoprotein function in Hs578T/Dox. Biochem Biophys Res Commun. 320:868–874. 2004.
Chen XP., Wang Q., Guan J., Huang ZY., Zhang WG., Zhang BX. Reversing multidrug resistance by RNA interference through the suppression of MDR1 gene in human hepatoma cells. World J Gastroenterol. 12:3332–3337. 2006.
Davidson B., Goldberg I., Givant-Horwitz V., Nesland JM., Berner A., Bryne M., Risberg B., Kopolovic J., Kristensen GB., Tropé CG., van de Putte G., Reich R. Caveolin-1 expression in ovarian carcinoma is MDR1 independent. Am J Clin Pathol. 117:225–234. 2002.
Dean M., Rzhetsky A., Allikmets R. The human ATP-binding cassette (ABC) t ransporter superfamily. Genome Res. 11:1156–1166. 2001.
Demeule M., Jodoin J., Gingras D., Beliveau R. P-glycoprotein is localized in caveolae in resistant cells and in brain capillaries. FEBS Lett. 466:219–224. 2000.
Farazi PA., DePinho RA. Hepatocellular carcinoma pathogenesis: from genes to environment. Nat Rev Cancer. 6:674–687. 2006.
Fielding CJ., Fielding PE. Caveolae and intracellular trafficking of cholesterol. Adv Drug Deliv Rev. 49:251–264. 2001.
Galbiati F., Volonte D., Meani D., Milligan G., Lublin DM., Lisanti MP., Parenti M. The dually acylated NH2-terminal domain of gi1alpha is sufficient to target a green fluorescent protein reporter to caveolin-enriched plasma membrane domains. Palmitoylation of caveolin-1 is required for the recognition of dually acylated g-protein alpha subunits in vivo. J Biol Chem. 274:5843–5850. 1999.
Gottesman MM., Fojo T., Bates SE. Multidrug resistance in cancer: role of ATP-dependent transporters. Nat Rev Cancer. 2:48–58. 2002.
Haber M., Burkhart CA., Regl DL., Madafiglio J., Norris MD., Horwitz SB. Altered expression of M beta 2, the class II beta-tubulin isotype, in a murine J774.2 cell line with a high level of taxol resistance. J Biol Chem. 270:31269–31275. 1995.
Higgins CF. ABC transporters: from microorganisms to man. Annu Rev Cell Biol. 8:67–113. 1992.
Kogo H., Fujimoto T. Caveolin-1 isoforms are encoded by distinct mRNAs. Identification of mouse caveolin-1 mRNA variants caused by alternative transcription initiation and splicing. FEBS Lett. 465:119–123. 2000.
Krajewska WM., Maslowska I. Caveolins: structure and function in signal transduction. Cell Mol Biol Lett. 9:195–220. 2004.
Lavie Y., Fiucci G., Liscovitch M. Up-regulation of caveolae and caveolar constituents in multidrug-resistant cancer cells. J Biol Chem. 273:32380–32383. 1998.
Lavie Y., Fiucci G., Liscovitch M. Upregulation of caveolin in multidrug resistant cancer cells: functional implications. Adv Drug Deliv Rev. 49:317–323. 2001.
Li L., Ren CH., Tahir SA., Ren C., Thompson TC. Caveolin-1 maintains activated Akt in prostate cancer cells through scaffolding domain binding site interactions with and inhibition of serine/threonine protein phosphatases PP1 and PP2A. Mol Cell Biol. 23:9389–9404. 2003.
Li S., Okamoto T., Chun M., Sargiacomo M., Casanova JE., Hansen SH., Nishimoto I., Lisanti MP. Evidence for a regulated interaction between heterotrimeric G proteins and caveolin. J Biol Chem. 270:15693–15701. 1995.
Li S., Song KS., Koh SS., Kikuchi A., Lisanti MP. Baculovirus-based expression of mammalian caveolin in Sf21 insect cells. A model system for the biochemical and morphological study of caveolae biogenesis. J Biol Chem. 271:28647–28654. 1996.
Liscovitch M., Lavie Y. Multidrug resistance: a role for cholesterol efflux pathways? Trends Biochem Sci. 25:530–534. 2000.
Liu J., Oh P., Horner T., Rogers RA., Schnitzer JE. Organized endothelial cell surface signal transduction in caveolae distinct from glycosylphosphatidylinositol-anchored protein microdomains. J Biol Chem. 272:7211–7222. 1997.
Liu P., Rudick M., Anderson RG. Multiple functions of caveolin-1. J Biol Chem. 277:41295–41298. 2002.
Nerenstone SR., Ihde DC., Friedman MA. Clinical trials in primary hepatocellular carcinoma: current status and future directions. Cancer Treat Rev. 15:1–31. 1988.
Nieth C., Priebsch A., Stege A., Lage H. Modulation of the classical multidrug resistance (MDR) phenotype by RNA interference (RNAi). FEBS Lett. 545:144–150. 2003.
Okamoto T., Schlegel A., Scherer PE., Lisanti MP. Caveolins, a family of scaffolding proteins for organizing “preassembled signaling complexes” at the plasma membrane. J Biol Chem. 273:5419–5422. 1998.
Pang A., Au WY., Kwong YL. Caveolin-1 gene is coordinately regulated with the multidrug resistance 1 gene in normal and leukemic bone marrow. Leuk Res. 28:973–977. 2004.
Quest AF., Leyton L., Parraga M. Caveolins, caveolae, and lipid rafts in cellular transport, signaling, and disease. Biochem Cell Biol. 82:129–144. 2004.
Razani B., Engelman JA., Wang XB., Schubert W., Zhang XL., Marks CB., Macaluso F., Russell R G. Li M, Pestell RG, Di Vizio D, Hou H Jr, Kneitz B, Lagaud G, Christ GJ, Edelmann W, Lisanti MP. Caveolin-1 null mice are viable but show evidence of hyperproliferative and vascular abnormalities. J Biol Chem. 276:38121–38138. 2001a.
Razani B., Zhang XL., Bitzer M., von Gersdorff G., Bottinger EP., Lisanti MP. Caveolin-1 regulates transforming growth factor (TGF)-beta/SMAD signaling through an interaction with the TGF-beta type I receptor. J Biol Chem. 276:6727–6738. 2001b.
Rosenbaum C., Rohrs S., Muller O., Waldmann H. Modulation of MRP-1-mediated multidrug resistance by indomethacin analogues. J Med Chem. 48:1179–1187. 2005.
Roy S., Luetterforst R., Harding A., Apolloni A., Etheridge M., Stang E., Rolls B., Hancock JF., Parton RG. Dominant-negative caveolin inhibits H-Ras function by disrupting cholesterol-rich plasma membrane domains. Nat Cell Biol. 1:98–105. 1999.
Schlegel A., Schwab RB., Scherer PE., Lisanti MP. A role for the caveolin scaffolding domain in mediating the membrane attachment of caveolin-1. The caveolin scaffolding domain is both necessary and sufficient for membrane binding in vitro. J Biol Chem. 274:22660–22667. 1999a.
Schlegel A., Wang C., Katzenellenbogen BS., Pestell RG., Lisanti MP. Caveolin-1 potentiates estrogen receptor alpha (ERalpha) signaling. caveolin-1 drives ligand-independent nuclear translocation and activation of ERalpha. J Biol Chem. 274:33551–33556. 1999b.
Scotto KW., Johnson RA. Transcription of the multidrug resistance gene MDR1: a therapeutic target. Mol Interv. 1:117–125. 2001.
Shatz M., Liscovitch M. Caveolin-1 and cancer multidrug resistance: coordinate regulation of pro-survival proteins? Leuk Res. 28:907–908. 2004.
Westermann M., Leutbecher H., Meyer HW. Membrane structure of caveolae and isolated caveolin-rich vesicles. Histochem Cell Biol. 111:71–81. 1999.
Williams TM., Lisanti MP. Caveolin-1 in oncogenic transformation, cancer, and metastasis. Am J Physiol Cell Physiol. 288:C494–506. 2005.
Yang CP., Galbiati F., Volonte D., Horwitz SB., Lisanti MP. Upregulation of caveolin-1 and caveolae organelles in Taxol-resistant A549 cells. FEBS Lett. 439:368–372. 1998.
Zhu H., Cai C., Chen J. Suppression of P-glycoprotein gene expression in Hs578T/Dox by the overexpression of caveolin-1. FEBS Lett. 576:369–374. 2004.
Go to :
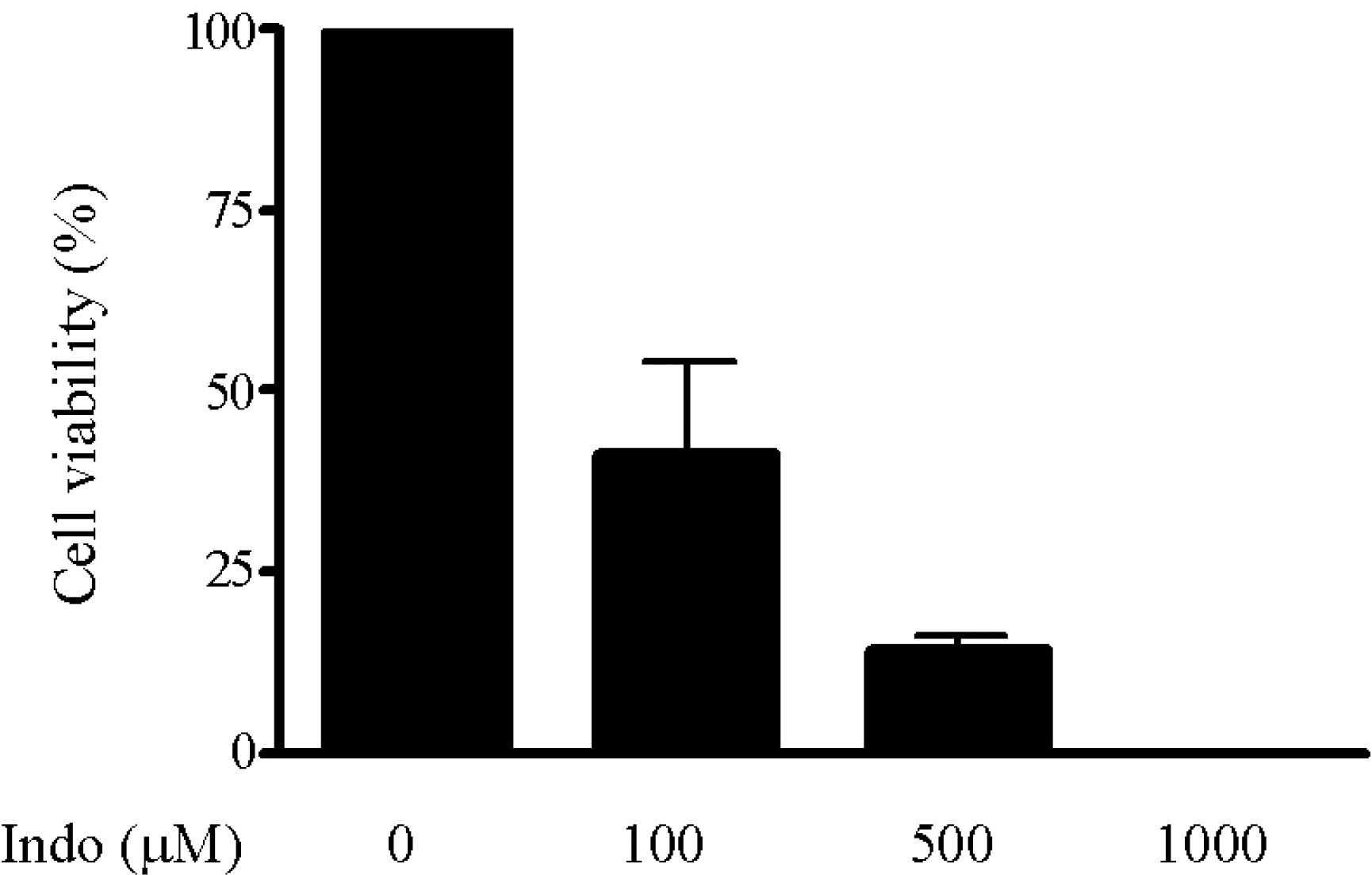 | Fig. 1.Indomethacin-induced cell death. Cell viability was determined by cell counting after treatment with different concentrations of indomethacin (0, 100, 500, and 1,000μM) for 24 h. |
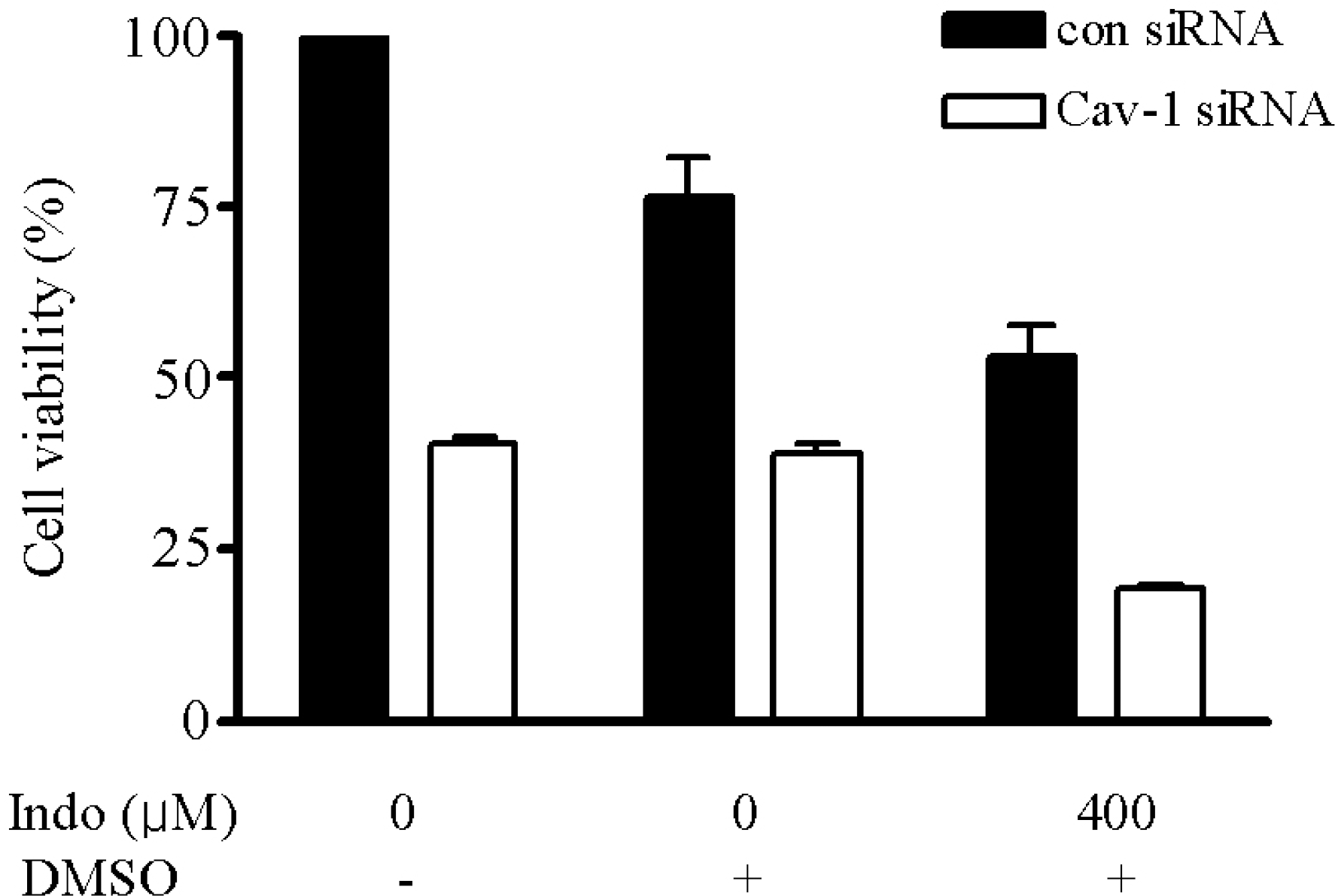 | Fig. 2.Viability of CAV1-siRNA-transfected cells. Cell viability was determined by cell counting after treatment with indomethacin (400μM) for 24 h. Cell viability was decreased significantly in CAV1-siRNA-transfected SK-Hep1 cells, and was markedly decreased by indomethacin treatment compared with that in each control. |
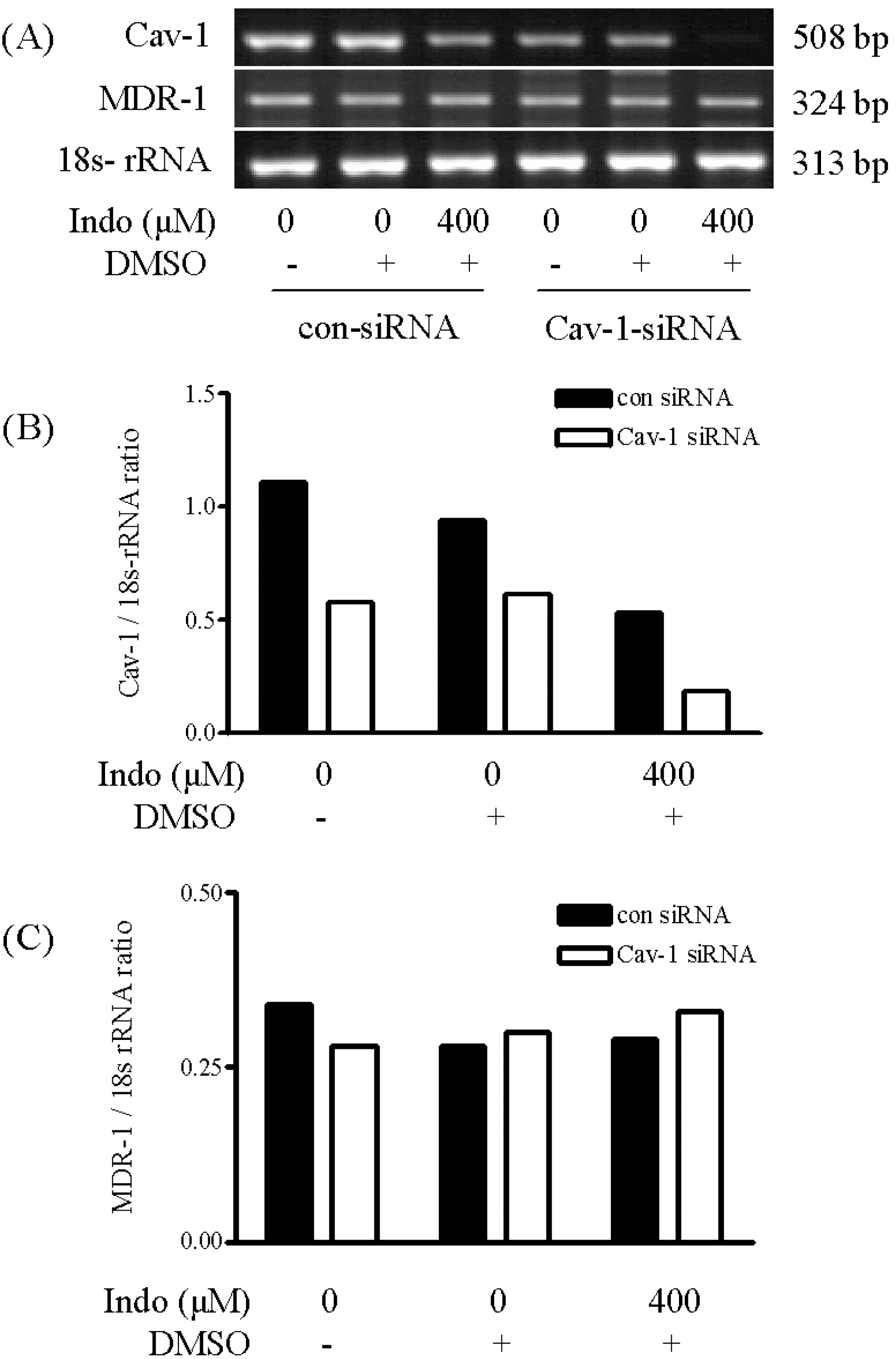 | Fig. 3.Effects of indomethacin treatment on the expression of CAV1 and MDR1 mRNAs in CAV1-siRNA-transfected cells. Cells were first transfected for 24 h with control siRNA or CAV1 siRNA. To complete the knockdown of CAV1 gene expression, a second transfection was performed for 24 h. Forty-eight hours after the first transfection, the cells were treated with indomethacin (400μ M) for 24 h. DMSO was used as the vehicle. Expression levels of CAV1 (B) and MDR1 (C) mRNAs were determined by RT-PCR (A) and normalized against that for 18S rRNA. |
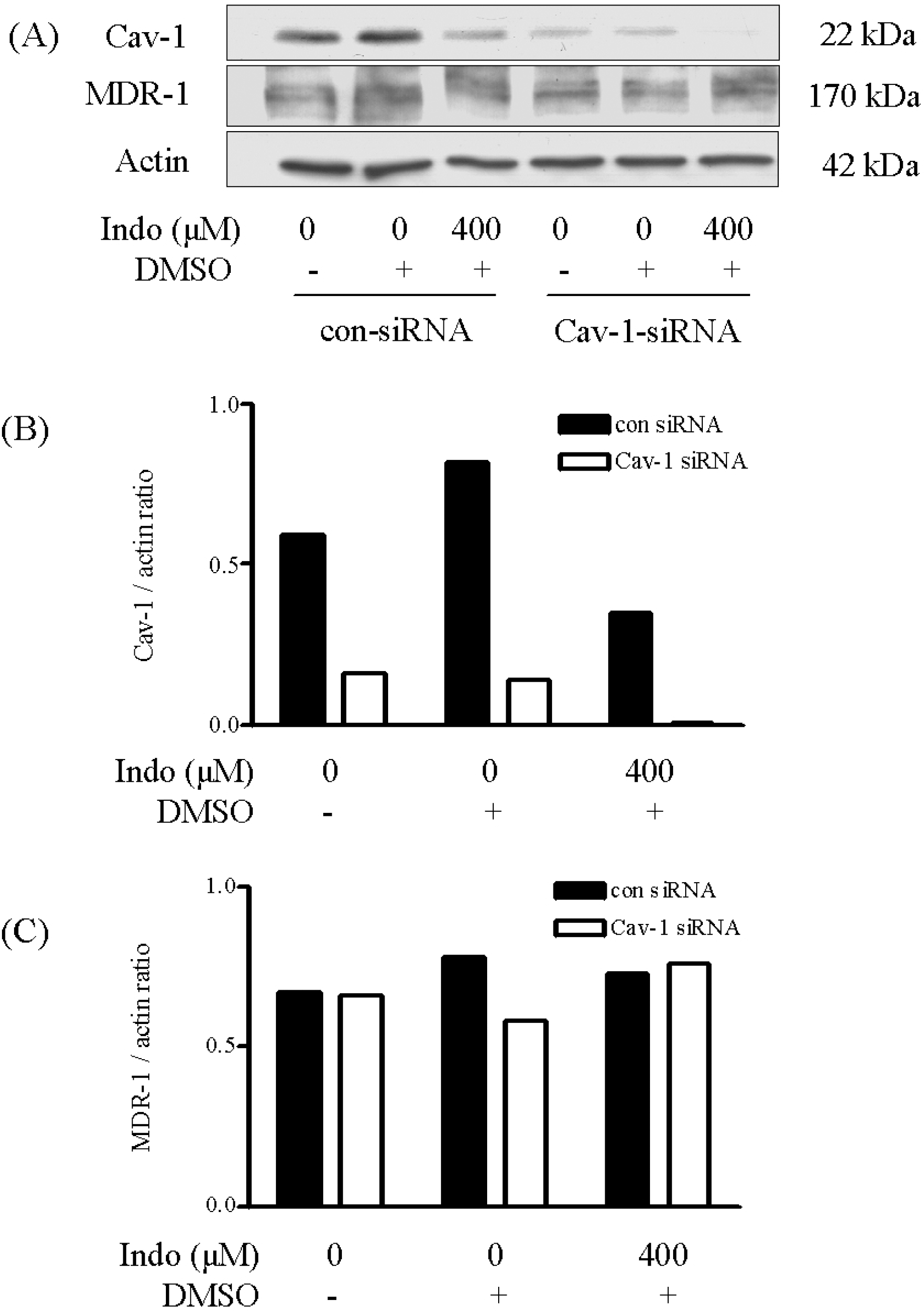 | Fig. 4.Effects of indomethacin treatment on the expression of CAV1 and MDR1 proteins in CAV1-siRNA-transfected cells. Crude protein was extracted from the cells. DMSO was used as the vehicle. Expression levels of CAV1 (B) and MDR1 (C) proteins were determined by Western-blot analysis (A) and normalized against those for β-actin. |
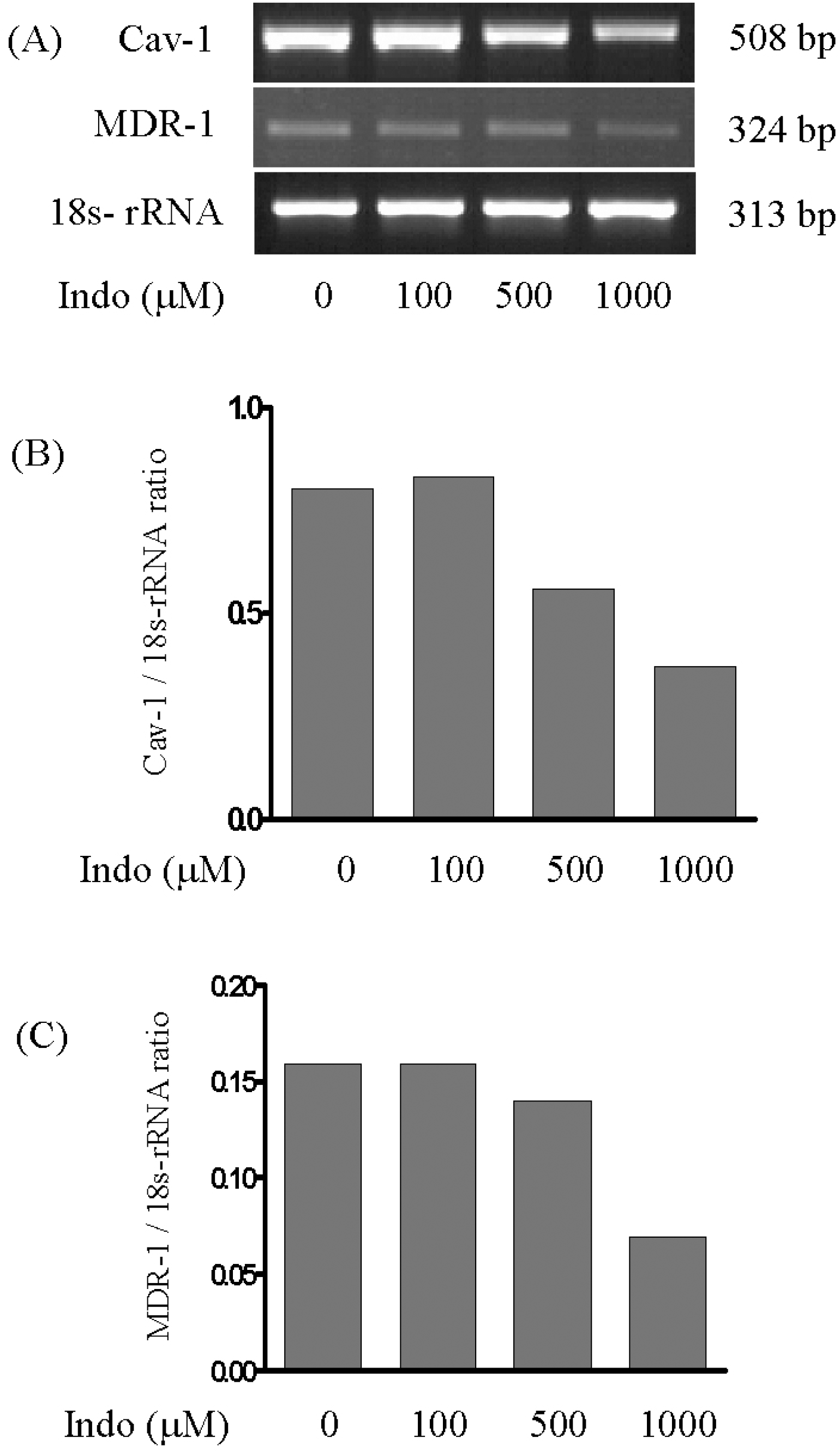 | Fig. 5.Effects of indomethacin treatment on the expression of CAV1 and MDR1 mRNAs in SK-Hep1 cells. Cells were treated with different concentrations of indomethacin (0, 100, 500, and 1,000μM) for 31 h. Expression levels of CAV1 (B) and MDR1 (C) mRNAs were determined by RT-PCR (A) and normalized against those for 18S rRNA. |
 | Fig. 6.Effects of indomethacin treatment on the expression of CAV1 and MDR1 proteins in SK-Hep1 cells. Crude protein was extracted from cells that had been treated with different concentrations of indomethacin (0, 100, 500, and 1,000μM) for 31 h. Expression levels of CAV1 and MDR1 proteins were determined by Westernblot analysis and normalized against those for β-actin. |
Table 1.
Nucleotide sequences of primers used in RT-PCR analysis
 XML Download
XML Download