

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Pentraxins are a superfamily of conserved proteins that are characterized by a cyclic multimeric structure and a conserved C-terminal domain. Classic pentraxins such as C-reactive protein and serum amyloid P are acute-phase proteins made in the liver, that are rapidly activated in response to inflammation [1]. Pentraxin 3 (PTX3, also called TNF-stimulated gene) is the prototypic long pentraxin. It is rapidly produced and released by several cell types, including endothelial cells, fibroblasts, retinal pigment epithelial cells, and particularly mononuclear phagocytes in response to either inflammatory or atheroprotective signals [2345].
PTX3 levels are very low in the serum and tissues of normal subjects; however, its expression rapidly increases in response to inflammatory stimulation in a wide range of diseases, including infectious, autoimmune, and degenerative disorders [678]. Specifically, PTX3 acts as a soluble pathogen recognition receptor with an essential role in resistance against selected pathogens [589]. Several studies have demonstrated that PTX3 is involved in the removal of apoptotic cells during immune response [101112]. PTX3 siRNA knockdown was also shown to promoted cell death, characterized as apoptosis and necrosis, in the presence of proinflammatory cytokines such as IL-1β and TNF-α in conjunctivochalasis fibroblasts [13].
The endoplasmic reticulum (ER) is a key site in the cell for protein folding and trafficking and is central to many cellular functions. Failure of the homeostatic capacity of the ER results in activation of the unfolded protein response (UPR), which leads to the elevated expression of ER chaperones and genes involved in ER expansion, as well as molecules affecting ER and cellular functions. Activation of the UPR generally reflects a loss of ER homeostasis, a condition referred to as ER stress. ER stress plays a fundamental role in the pathogenesis of several diseases such as diabetes, cancer, and neurodegenerative diseases. In ophthalmology, diseases such as glaucoma, diabetic retinopathy, and age-related macular degeneration (ARMD) display features characteristic of ER stress.
In higher eukaryotes, UPR signaling is initiated by three ER transmembrane sensors: inositol-requiring enzyme 1α (IRE1α), pancreatic ER kinase-like ER kinase (PERK), and activating transcription factor 6 [1415]. If proper protein folding capacity in the ER cannot be restored, the UPR upregulates genes such as CCAAT-enhancer-binding protein homologous protein (CHOP), which activates apoptotic pathways [16].
ARMD is a multi-factorial disease and a leading cause of visual impairment in the elderly. Recent studies suggest that production of reactive oxygen species and chronic oxidative stress may play a pivotal role in the development of this disease. The retina, especially the retinal pigment epithelium (RPE), is exposed to high levels of oxidative and photo-oxidative damage over a lifetime. Oxidative stress can inactivate chaperones, promote aberrant disulfide bond formation, promote stabilization of undesirable intermediates, and inhibit degradation of misfolded proteins, which together cause ER stress. However, recent genetic studies suggest a role in the immune response, particularly abnormalities in the innate immune system, in the pathogenesis and severity of ARMD.
The RPE plays an important role in establishing the immune privilege of the eye by secreting immunosuppressive factors [1718]. With these complex and varied roles, the RPE is essential for visual function. Failure in any one of its functions can lead to degeneration of the retina, loss of visual function, and blindness. The RPE, which is the major target cell of ARMD, may be a site for crosstalk between cellular oxidative ER stress and immune signaling. PTX3 produced in RPE cells may play an important protective role in retinal injury against proapoptotic stimuli such as ER stress. In this study, we examined whether PTX3 is regulated by tunicamycin-induced ER stress. We also investigated the role of PTX3 in ER stress-associated cell death in the human retinal pigment epithelial cell line, ARPE-19.
Materials and Methods
Cell culture
ARPE-19 cells (Fig. 1A and 1B), a human retinal pigment epithelial cell line, were purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA). They were cultured in T-75 flasks with Dulbecco's modified Eagle's medium (Invitrogen, Gibco, Carlsbad, CA, USA) supplemented with 10% fetal bovine serum (Sigma-Aldrich, St. Louis, MO, USA) and 100 U/mL penicillin and streptomycin (Gibco-BRL, Gaithersburg, MD, USA). During incubation, the culture medium was changed every 2 days. All cultures were maintained at 37℃ under 5% CO2 with 95% relative humidity.
Antibodies and reagents
Protein kinase inhibitors such as NF-κB inhibitor (Bay 11-7085) and a c-Jun NH2-terminal kinase (JNK) MAP kinase inhibitor (SP600125; Enzo Life Sciences, Montgomery County, PA, USA), human TNF-α (R&D Systems, Minneapolis, MN, USA), human IL-1β (Abcam, Cambridge, MA, USA), and human IFN-γ (PeproTech, Rocky Hill, NJ, USA) were used. Human PTX3 monoclonal antibody (Novus Biologicals, Littleton, CO, USA) and β-actin polyclonal antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA) were used for western blot analysis. All signaling antibodies were purchased from Cell Signaling Technology (Danvers, MA, USA).
RNA isolation and real-time reverse transcription-polymerase chain reaction
Total RNA from ARPE-19 cells was isolated using a reagent (Invitrogen), and reverse transcription was performed using Moloney murine leukemia virus reverse transcriptase (Invitrogen). The amplification of cDNA by polymerase chain reaction was performed using human PTX3-specific primers and the AccuPower PCR PreMix kit (Bioneer, Daejeon, Korea). β-actin mRNA expression served as a control. Primers were designed using the Primer Express 1.5 software (Applied Biosystems, Carlsbad, CA, USA) and synthesized by Bioneer. The primer sequences were as follows: human PTX3 forward primer, 5′-AATGCATCTCCTTGCGATTC-3′; reverse primer, 5′-TGAAGTGCTTGTCCCATTCC-3′; IRE1α forward primer, 5′-CACAGTGAC GCTTCCTGAAAC-3′; reverse primer, 5′-GCCATCATTAGGATCTGGGAGA-3′; CHOP forward primer, 5′-CCGTTTCCTGGTTCTCCCTTGG-3′; reverse primer, 5′-TGCTTCTCT GGCTTGGCTGAC-3′, and β-actin forward primer, 5′-ATGGTGCGTGACATTAAGGAGAAG-3′; reverse primer, 5′-AGGAAGGAAGGCTGGAAGAGTG-3′. Amplification of cDNA started with 10 minutes at 95℃, followed by 40 cycles of 15 seconds at 95℃ and 1 minute at 59℃. Reverse transcription-polymerase chain reaction for PTX3 ; IRE1α, CHOP, and β-actin was conducted using iQ SYBR Green Supermix (Bio-Rad, Hercules, CA, USA).
Western blot analysis
Western blot analyses were performed as previously described [19]. Briefly, the cells were harvested using RIPA buffer (Sigma-Aldrich) containing protease inhibitors (Roche Applied Science, Mannheim, Germany). Protein concentrations were determined using the Pierce BCA protein assay kit (Thermo Scientific, Rockford, IL, USA). Samples were resolved by 12% SDS-PAGE gels and transferred to PVDF membranes (Bio-Rad) overnight (120 mA). The transferred membranes were hybridized with various primary antibodies (diluted 1 : 1000, Cell Signaling Technology) overnight at 4℃, followed by incubation with secondary HRP-conjugated IgG antibodies and visualized with SuperSignal West Pico Chemiluminescent Substrate (Thermo Scientific).
Enzyme-linked immunosorbent assay
Human PTX3 released into the culture medium was measured using an enzyme-linked immunosorbent assay (ELISA) kit (human PTX3/TSG-14) from R&D Systems, according to the manufacturer's instructions. Briefly, ELISA plates (BD Biosciences, San Jose, CA, USA) were coated with a monoclonal anti-human PTX3 antibody (2 µg/mL) in coating buffer (1% BSA in PBS, pH 7.2 to 7.4) overnight at room temperature. The plates were then blocked with coating buffer for 2 hours at room temperature and incubated with either recombinant human PTX3 standards or samples collected in quadruplicate (100 µL/well) for another 2 hours. The plates were then incubated with a biotinylated human PTX3 antibody (150 ng/mL) for 2 hours and subsequently incubated with freshly diluted streptavidin-HRP for 20 minutes in the dark. After each step, the plates were washed three times with washing buffer. A chromogenic substrate tetramethylbenzidine (100 µL/well) (eBioscience, San Diego, CA, USA) was added and incubated for 5 minutes in the dark. The reaction was stopped by adding 2 N H2SO4 (50 µL/well), and the plates were read at 450 nm using an automatic ELISA reader (Merck Sensident Scan, Helsinki, Finland)
Cell viability assay
Cell viability was determined by the MTS assay using the CellTiter 96 AQueous one solution cell proliferation assay kit (Promega, Madison, WI, USA). Cells were seeded at 0.7 × 104 cells per well in 96-well plates. After reagent treatment, 20 µL of MTS solution was added to each well, and plates were incubated for an additional 2 to 4 hours at 37℃. Absorbance was then measured at 490 nm using a SpectraMax M2 microplate reader (Molecular Devices, Sunnyvale, CA, USA) to calculate cell survival percentages.
Nuclear staining
Cells were seeded on 12-well plates containing glass coverslips and treated the following day with tunicamycin (1 µM) for 48 hours. Nuclear counterstaining was performed by staining with a solution of 1 µg/mL Hoechst 33258 for 5 minutes and mounting on a slide using fluorescence mounting medium (Dako, Glostrup, Denmark). Cell viability was assessed using a fluorescence microscope.
Small hairpin RNA knockdown of IRE1α and PTX3
ARPE-19 cells stably transfected with the pMSCV-LMP retro-puro vector carrying specific small hairpin RNA (shRNA) targeting nucleotides to PTX3 (PTX3 shRNA) or IRE1α (IRE1α shRNA) were generated according to the manufacturer's instructions (OPEN Biosystems, Lafayette, CO, USA). Briefly, the PTX3 shRNA oligo (5′-TGCTGTTGACAGTGAGCGCCCATCAACCTCTCTTCTGTATAGTGAAGCCACAGATGTATACAGAAGAGAGGTTGATGGGTGCCTACTGCCTCGGA-3′) and IRE1α shRNA oligo (5′-TGCTGTTGACAGTGAGCGCAGTGCATAATAGGAACACTTTAGTGAAGCCACAGATGTAAAGTGTTCCTATTATGCACTGTGCCTACTGCCTCGGA-3′) were synthesized and amplified using miR30 common primers (forward primer: 5′-CAGAAGGCTCGAGAAGGTATATTGCTGTTGACAGTGAGCG-3′, reverse primer: 5′-CTAAAGTAGCCCCTTGAATTCCGAGGCAGTAGGCA-3′). The polymerase chain reaction products were digested with EcoRI and XhoI and ligated into pMSCV-LMP retro-puro (OPEN Biosystems). Transfected cells were selected in media containing 2 µg/mL puromycin (Life technologies, Carlsbad, CA, USA).
Statistical analysis
Data are represented as mean ± standard deviation. For comparisons between two groups, we used the Student's two-tailed unpaired t-test. For comparisons of timed series experiments, we performed Student's paired t-tests. Statistically significant differences were designated at p < 0.05.
Results
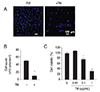
ER stress is linked to several pathological conditions, including ARMD [202122]. To verify the cellular effects of the ER stress inducer, tunicamycin, ARPE-19 cells were exposed to 1 µg/mL tunicamycin or the vehicle control (1% DMSO) for 48 hours. The cells were harvested and nuclei stained using Hoechst 33342 (Fig. 2A). Live cells were counted and are represented as a graph in Fig. 2B and 2C. Tunicamycin-induced ER stress accelerated the death of ARPE-19 cells. To verify these results, we assessed ER stress-induced cell death in the absence or presence of tunicamycin using the CellTiter AQueous one solution assay kit (Promega, Leiden, The Netherlands). The viability of ARPE-19 cells decreased with 1 µg/mL tunicamycin treatment (24.2%) compared to vehicle (100%). These results indicate that excessive ER stress induces cell death in ARPE-19 cells [2223].
To investigate the effects of ER stress on PTX3 production, PTX3 expression was measured from supernatants after 48-hour treatment with the vehicle or tunicamycin (0.01, 0.1, and 1 µg/mL) (Fig. 3A). Tunicamycin induced PTX3 production in a dose-dependent manner. Furthermore, a time course analysis in which ARPE-19 cells were treated with 1 µg/mL tunicamycin for 6, 12, 24, 36, and 48 hours demonstrated that PTX3 production began to increase by 12 hours, and a more striking increase in PTX3 was evident after 36 hours of tunicamycin treatment (Fig. 3B). PTX3 production by tunicamycin treatment was significantly enhanced by 48 hours. These results suggest that tunicamycin induced ER stress can enhance the production of PTX3 in human retinal pigment epithelial cells in both a dose and time dependent manner.
To ascertain the effect of tunicamycin induced ER stress on PTX3 mRNA levels in ARPE-19 cells, total RNA was isolated 24 hours after treatment with increasing doses of tunicamycin (0.01, 0.1, and 1 µg/mL) and at different time points (3, 6, 12, 24, 36, and 48 hours) using 0.1 µg/mL of tunicamycin (Fig. 4A and 4B). PTX3 mRNA levels increased with increasing tunicamycin concentration (Fig. 4A). Furthermore, PTX3 mRNA began to increase by 6 hours, and a more striking increase was observed after 36 hours of tunicamycin treatment (Fig. 4B). Taken together, these results suggest that tunicamycin-induced ER stress increased PTX3 production at the transcriptional level.
In order to verify which signaling pathway could be responsible for the stimulation of PTX3 production by tunicamycin-induced ER stress, we used a specific NF-κB inhibitor (Bay 11-7085) and a JNK MAP kinase inhibitor (SP600125). ARPE-19 cells were treated with Bay 11-7085 (10 µM) and SP600125 (10 µM) in the presence of tunicamycin, and supernatants were harvested 48 hours after treatment. Bay 11-7085 blocked the stimulation of PTX3 production by tunicamycin (Fig. 5A); however, SP600125 had no effect on PTX3 production in the presence of tunicamycin (Fig. 5A). These data suggest that the NF-κB signaling pathway may facilitate increased PTX3 production during ER stress in human retinal pigment epithelial cells.
In higher eukaryotes, there are three major ER stress sensors, IRE1α, PERK, and activating transcription factor 6 [2425]. These three ER stress response branches control the expression of specific transcription factors and signaling events that modulate a variety of downstream ER stress responses, which orchestrate adaptation to ER stress. In addition to upregulating UPR target genes, mammalian IRE1α signals through additional pathways include NF-κB and JNK [26]. To determine whether IRE1α is involved in the upregulation of PTX3 during ER stress, we generated control and IRE1α shRNA transfected ARPE-19 cells. Real-time reverse transcription-polymerase chain reaction analyses were performed to assess the degree of IRE1α silencing (Fig. 5B). IRE1α, shRNA transfected ARPE-19 cells abrogated tunicamycin-induced PTX3 production (Fig. 5C). These data suggest that IRE1α is an important mediator of PTX3 production by the ER stress inducer, tunicamycin.
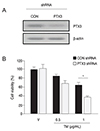
To investigate whether PTX3 production altered cell viability during ER stress, we generated control and PTX3 shRNA-transfected ARPE-19 cells. Western blot analyses were performed to assess the degree of PTX3 silencing (Fig. 6A). The cell viability of PTX3 shRNA transfected ARPE-19 cells decreased with tunicamycin treatment (1 µg/mL, 30.8%), as compared to the vehicle-treated control (55.4%) (Fig. 6B). These data suggest that PTX3 promotes cell survival during ER stress induced by tunicamycin in ARPE-19 cells.
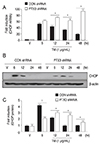
During physiological ER stress, transient CHOP expression is beneficial. However, in pathologically chronic ER stress, prolonged CHOP expression promotes cell death, and it has been implicated in a number of diseases, including ARMD, neurodegenerative diseases, atherosclerosis, diabetes, and renal disease [27282930]. To investigate the effects of PTX3 knockdown on CHOP expression, we treated control and PTX3 shRNA transfected ARPE-19 cells with tunicamycin (1 µg/mL) and harvested RNA and protein at 6, 12, 24, and 48 hours after treatment. CHOP mRNA and protein levels increased after 6 hours and declined at 12 hours in the control shRNA transfected ARPE-19 cells (Fig. 7A and 7B, respectively). However, CHOP mRNA and protein levels increased after 6 hours, and they were prolonged until 48 hours (mRNA) or 24 hours (protein) in PTX3 shRNA-transfected ARPE-19 cells (Fig. 7A and 7B, respectively). Fold changes in the signal intensity of protein expression were calculated in comparison to the control in control shRNA transfected cells (black) or PTX3 shRNA transfected cells (white) (Fig. 7C). Taken together, these results indicate that ER stress stimulates PTX3 production via an IRE1α-dependent signaling pathway that promotes cell survival in response to ER stress-induced cell death in ARPE-19 cells.
Discussion
RPE cell absorb light energy focused on the retina by the lens, and contribute to the maintenance of photoreceptor excitability. Damage to any of the intricate structures of the retina can lead to visual impairment and blindness [3132]. Apoptosis is a common form of cell death observed in RPE cells. It is also an important feature of ARMD, the most common cause of irreversible vision loss, particularly in individuals over 65 years of age [33]. Age-related alterations in the RPE include a reduction in cell density, which can be caused by apoptosis resulting from accumulation of toxic substances [34]. Oxidative stress, hyperglycemia, mitochondrial dysfunction, and ER stress are among the many studied pro-apoptotic factors in RPE cell lines [35363738]. Recent studies suggest that the ER may also play an important role in response to oxidative stress-induced damage in neuronal and epithelial cells [394041]. ER stress has been identified in the tissues of patients with inherited macular degeneration. Therefore, ER stress in RPE could play an important role in the pathogenesis of choroidal neovascularization complicating AMD [4243].
Pentraxins are key components of the humoral arm of innate immunity, which also include the complement system, collectins, and ficolins [1]. PTX3 is the first member of the long pentraxin subfamily and was identified in the early 1990s as a new secreted protein rapidly induced by IL-1β in endothelial cells or by TNF in fibroblasts [4445]. PTX3 plays a non-redundant role in humoral innate immunity, assisting in the resistance to selected pathogens by recognizing microbes, activating complement, and facilitating pathogen recognition by phagocytes. PTX3 is also essential in female fertility [446474849] and acts as a scavenger of cell debris, as well as an immunosuppressant. It specifically binds to late apoptotic cells and subsequently inhibits their uptake by dendritic cells, thus acting as an additional tool for maintaining immune tolerance [1050]. PTX3 binds to complement factor H, a complement regulator protein whose main function is to inhibit activation of the alternative complement pathway. Its polymorphic variant, harboring a substitution of histidine for tyrosine at codon Y420H, has been shown to be strongly associated with ARMD [51].
We previously reported that human retinal pigment epithelial cells are a major source for PTX3 production in the presence of the proinflammatory cytokines IL-1β and TNF-α, and they could be important mediators for host defense and inflammatory response in the retina [52]. We also showed that PTX3 concentration was significantly higher in neovascular ARMD patients and diabetic retinopathy patients than in control patients [5354]. These data suggest that PTX3 produced in RPE cells may play an important role in protection against inflammation and proapoptotic factors, including ER stress, during retinal injury. Furthermore, Wu et al. [55] demonstrated that PTX-3 secretion was regulated during thapsigargin-induced ER stress in cumulus-oocyte complexes.
In this study, we found that a human RPE cell line, ARPE-19, locally produced PTX3, and its expression and production were increased by tunicamycin-induced ER stress via an IRE1α-mediated NF-κB signaling pathway. As shown in Fig. 6, PTX3 shRNA transfected cells were more sensitive to tunicamycin-induced cell death. This was because of prolonged expression of CHOP, a mediator of apoptosis during ER stress (Fig. 7). These data suggest that PTX3 produced in RPE cells play an important protective role in retinal injury against proapoptotic stimuli such as ER stress. The role of PTX3 production in the presence of other pathological stimuli related to retinal degenerative and other eye diseases needs to be investigated.




 XML Download
XML Download