

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Hypoxia is one of the leading causes of retinal vascular pathology and neuronal cell damage, which are commonly observed in retinal pathologies such as retinopathy of prematurity (ROP) and diabetic retinopathy [1,2]. Oxygen-induced retinopathy (OIR) in neonatal rats, under conditions where neuronal damage is induced by hypoxic retinal damage, is a widely used animal model of ROP [3-5]. Kim et al. [6] reported that the most severe retinal injury from hyperoxic/normoxic injury appears in OIR rat retinas at postnatal day 18 (P18) and 4 days after relative hypoxia (from P2 to P14).
Under certain pathological conditions, expression of neuronal and inducible nitric oxide synthase (nNOS and iNOS, respectively) might be enhanced, resulting in neurotoxicity and subsequent neurodegeneration [7,8]. Kaur et al. [9] and Kaur et al. [10] suggested that increased vascular endothelial growth factor (VEGF) and nitric oxide (NO) production in hypoxia results in increased vascular permeability and degeneration of neural cells. The mechanism leading to excessive nNOS activation is thought to be mediated by hyperactivation of glutamate receptors, as well as possible mitochondrial dysfunction and cell death [11]. In addition, NO is postulated as a key neurotoxic factor of ROP and an increase in NO concentration in retina induces cytotoxic effects [12-15].
Triamcinolone acetonide (TA) is a corticosteroid with anti-inflammatory and anti-angiogenic effects that has been reported to influence cellular events in vascular and neural tissues through the activation of cytokines [9,10,16-18]. Various reports have suggested that TA may have a neuroprotective effect in retinal disorders and may be involved in neuroprotection of neuronal and glial cells [19,20]. In a model of OIR, Hartnett et al. [21] suggested that TA reduces neovascularization and capillary density, while Park et al. [22] reported that TA protects retinal neurons damaged by relative hypoxia from decreased decorin levels.
In this study, we describe the effect of TA treatment on the expression of nNOS and iNOS in the retinas of OIR rats.
Materials and Methods
Animals
Pregnant Sprague-Dawley (SD) rats were purchased from KOATEC (Pyeongtaek, Korea). Rats were individually housed under alternating 12-hour light/dark cycles and given rat chow and water, in accordance Use Committee of Gyeongsang National University.
Oxygen-induced retinopathy rat model
For promotion of OIR, five new born SD rat pups were exposed to hyperoxia (80 ± 1.3% O2) from P2 to P14 (with 2 hr/day in room air), and then returned to normal conditions (room air, 21 ± 1.5% O2) from P14 to P18, as previously described [6]. Five control rats were maintained in room air. All rats were sacrificed at P18 to investigate changes in OIR rat retinas compared with normal retinas.
Triamcinolone acetonide treatment
All rats were anesthetized at P15 to assess the effect of TA (Hanall Co., Seoul, Korea) on the retinas of OIR and control rats. Using a 30-gauge needle, a TA solution (2 µL, 40 mg/mL stock solution in saline) was injected under an operating microscope through the pars plana into the right vitreous, and a saline solution (2 µL) was injected into the left vitreous.
Total protein extraction and Western blot analysis
Total protein extraction and Western blot analysis was performed as previously described [6]. Retinal protein isolated from five OIR and five control rats at P18 with TA or saline treatment were subjected to 10% sodium dodecyl sulphate polyacrylamide gel electrophoresis and transferred to a nitrocellulose membrane. The blots were incubated in mouse monoclonal anti-nNOS antibody (1 : 1,000 dilution; BD Biosciences, San Jose, CA, USA) and in rabbit polyclonal anti-iNOS antibodies (1 : 500 dilution; Abcam, Cambridge, UK), followed by incubation with horseradish peroxidase-conjugated rabbit anti-mouse and goat anti-rabbit IgGs (Pierce, Rockford, IL, USA). Specific proteins were visualized using an enhanced chemiluminescent kit according to the manufacturer's instructions (Amersham Bioscience, Piscataway, NJ, USA). Blots were re-probed with an α-tubulin antibody (1 : 20,000 dilution; Sigma, St. Louis, MO, USA) as a control for loading. Changes in expression of retinal nNOS and iNOS protein were measured in five independent tests.
Tissue preparation and immunohistochemistry
Immunohistochemistry was performed on 12-µm retinal frozen sections from P18 OIR and control rats, as previously described [6]. Sections were blocked in 1.5% normal goat serum for 30 min at room temperature (RT), incubated with primary nNOS and iNOS antibodies (1 : 200 dilution, respectively) for 2 hours at 4℃, washed three times with 10 mmol/L Tris-buffered saline, and incubated with biotinylated anti-rabbit IgG antibody (1 : 200; Santa Cruz Biotechnology, Santa Cruz, CA, USA) in Tris buffer for 1 hour at RT. Following an additional wash with Tris buffer, sections were incubated with avidin-biotinylated horseradish peroxidase complex (ABC; Vector Laboratories Inc., Burlingame, CA, USA) and treated with 0.025% 3,3'-diaminobenzidine tetrahydrochloride (Sigma) in 0.003% H2O2.
Double immunofluorescence was performed in the following manner. After washing with Tris buffer, the sections were then incubated at RT with a cocktail of two primary antibodies, anti-nNOS and anti-parvalbumin as markers for amacrine cells, and iNOS and GFAP as markers for astrocytes. For immunofluorescent staining, Alexa Fluor-488 or -594 goat anti-rabbit or -mouse IgG were employed as secondary antibodies (1 : 200; Pierce; Invitrogen, Carlsbad, CA, USA). After several washes with Tris buffer, the sections were mounted using ProLong Gold antifade reagent (Invitrogen).
Imaging and statistical analyses
Retinal images were captured at a distance of approximately 0.8 to 1.0 mm from the optic nerve head using an IX2-DSU disk scanning confocal microscope (Olympus, Wendenstrasse, Hamburg, Germany). The number of cells that had a immunoreactive nNOS signal was quantified in the ganglion cell layer (GCL) and inner nuclear layer (INL) in whole fields containing five different retinas of saline or TA treated control and OIR rats group. Each experiment was independently performed five times using five different retinal samples for each group.
Densitometry measurements were processed using SigmaGel ver. 1.0 (Jandel Scientific, Erkrath, Germany) and SigmaPlot ver. 4.0 (SPSS Inc., Chicago, IL, USA). The significance of inter-group differences was evaluated after α-tubulin normalization in each group by Kruskal Wallis H-test and Mann-Whitney U-testing (SPSS, SPSS Inc.). Data are presented as mean ± SE. Differences were considered significant at p < 0.05.
Results
Effect of treatment with TA on retinal levels of nNOS and iNOS protein
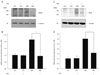
nNOS protein appeared as a single band of approximately 155 kDa when subjected to electrophoresis (Fig. 1A). We found increased retinal levels of nNOS protein at P18 in saline injected OIR rats (2.12-fold, p = 0.00134, n = 5) (Fig. 1B) compared with saline injected control rats. Once again, treatment with TA prevented the increase in retinal nNOS protein in OIR rats compared with saline injected control rats (1.00-fold, p = 0.625, n = 5) (Fig. 1B). The difference in expression of nNOS protein expression between TA and saline treated P18 rats was significant (p = 0.00057) (Fig. 1B).
iNOS protein appeared as a single band of approximately 130 kDa (Fig. 1C). We found increased retinal levels of iNOS protein at P18 in saline injected OIR rats (2.99-fold, p = 0.0027, n = 5) (Fig. 1D) compared with saline injected control rats. Treatment with TA prevented increased abundance of retinal iNOS protein in OIR rats compared with saline injected control rats (1.58-fold, p = 0.1371, n=5) (Fig. 1D). In addition, the difference in expression of iNOS protein in P18 OIR rats with or without TA treatment was significant (p = 0.011) (Fig. 1D).
Effect of treatment with TA on retinal immunoreactivity of nNOS and iNOS
Retinal nNOS immunoreacitvity was observed in neuronal cells, especially amacrine cells, in the GCL and INL of retinas from saline injected control rats (Fig. 2A). Increased nNOS immunoreactivity was more prevalent in the GCL and INL of retinas of saline injected OIR rats compared with saline injected control rats at P18 (Fig. 2B). Treatment with TA in OIR rats led to nNOS immunoreactivity similar to that observed in the retinas of saline injected control rats (Fig. 2C). Double staining revealed an overlap of nNOS signal (green) and the parvalbumin signal (red) in the GCL and INL of retinas of saline injected OIR rats (Fig. 2D). The nNOS-immunoreactive cells in the inner retina were determined to be parvalbumin-positive amacrine cells. The number of nNOS-positive amacrine cells in saline injected P18 OIR rats was significantly increased versus saline injected control rats (2.05-fold, p = 0.00001, n = 5) (Fig. 3A). The increase of nNOS-positive amacrine cells in P18 OIR rats was blocked by treatment of TA compared with saline injected control rats (1.43-fold, p = 0.0088, n = 5) (Fig. 3A). Furthermore, the difference in the number of nNOS-positive amacrine cells in P18 OIR rats between groups with and without treatment was significant (p = 0.00385) (Fig. 3A).
Weak iNOS expression was detected in the nerve fiber layer (NFL) of retinas from saline injected control rats (Fig. 2D). Increased iNOS immunoreactivity was observed in the NFL of saline injected OIR rats compared with saline injected control rats at P18 (Fig. 2F). Treatment with TA prevented the increase of nNOS immunoreactivity in OIR rats. Expression levels were similar to that seen in the retinas of saline injected control rats (Fig. 2G). Double immunofluorescent staining revealed an overlap of the iNOS signal (red) and the GFAP signal (green) in the NFL of saline injected OIR rats (Fig. 2H). The iNOS-immunoreactive cells observed in the inner retina were determined to be GFAP-positive astrocytes. The iNOS immunoreactivity in saline injected P18 OIR rats was significantly increased versus saline injected control rats (2.57-fold, p = 0.0012, n = 5) (Fig. 3B). The increase of iNOS expression in P18 OIR rats was blocked by treatment of TA compared with saline injected control rats (1.22-fold, p = 0.1173, n = 5) (Fig. 3B). In addition, the difference in iNOS expression in P18 OIR rats between rats with and without TA treatment was significant (p = 0.00035) (Fig. 3B).
Discussion
In this study, we examined the expression of nNOS and iNOS in the retinas of OIR rats. In previous studies, the expression of nNOS was found to increase in rat models of chronic glaucoma, diabetic retinopathy, and optic nerve transaction [16,23-25]. Other reports have suggested that the protein expression of eNOS and nNOS is increased in the OIR model, while the expression of iNOS is decreased [26]. This discrepancy is thought due to differences of experimental OIR models, especially the distribution of hyperoxia and length of normoxia period. In the present study, significant increases were observed for the expression of nNOS and iNOS in the OIR rat retina compared with control rats. Thus, under hypoxic conditions, nNOS and iNOS activation might be increased, leading to cellular changes that may play a role in neurodegeneration.
The developing eye and visual system have been reported to be susceptible to hypoxic-ischemic damage [27]. Oxygen deprivation induces overproduction of NO, an excess of which is toxic to cells [28,29]. Given that, at the retina, neurotoxicity induced by hypoxia is related to alterations in nitric oxide regulation [12], it may be suggested that the increased expression of iNOS and nNOS in the OIR retinas observed in our study might be responsible for neuronal damage.
Recently, Rey-Funes et al. [15] reported that hypothermia is protective against damage of the retina induced by perinatal asphyxia, which was due to inhibition of alterations of both NOS isoforms and nitration of proteins. Several strategies such as the use of antioxidant and dexamethasone have been tried in order to prevent hypoxia-ischemic injury [30,31]. Previously, we found retinal vascular pathologic changes and gene expression changes in OIR rats at P18 and demonstrated suppression of these changes via interruption of VEGF activation through exposure to TA [6,31,32]. In the present study, TA blocked an increase of iNOS and nNOS expression, suggesting that TA may be protective against damage of retina in OIR rat retinas. However, the mechanism by which treatment with TA prevents increased nNOS and iNOS expression has not been defined. Additional experiments will be required to determine whether TA treatment may affect neuronal cell damage and regeneration through the activation/inhibition of various cytokines in the hypoxic retina.
We analyzed a number of nNOS-positive cells in the INL and the GCL. The increase in the number of nNOS-positive amacrine cells in the INL and the GCL was determined in the retinas of OIR rats. In the retinas of normal rats, nNOS immunoreactivity was localized to certain populations of amacrine cells found in the INL and in a small group of neurons localized in the GCL [33]. These nNOS-containing neurons exhibited their own basal expression levels of nNOS in the retinal layers, and thus these neurons might be expected to react differentially in response to retinal ischemia [34]. Oh et al. [35] observed that nNOS-expressed neurons in normal rats co-localizes with gamma aminobutyric acid (GABA), suggesting that an increase of nNOS-expressing amacrine cells might be accompanied by dysfunction of GABA metabolism in diabetic retina. In addition, Rey-Funes et al. [15] demonstrated colocalizaion of nNOS and tyrosine hydroxylase (TH) in the cytoplasm of a subpopulation of ganglion and amacrine cells of perinatal asphyxia model rats, which could be prevented by hypothermia. However, in our study, nNOS-positive cells were co-localized with parvalbumin but not in GABA, choline acetyltransferase or TH (data not shown) expressing cells, future studies will be needed to determine the reason for this discrepancy. Otherwise, the present study using double immunofluorescence labeling with GFAP (a marker for astrocytes) and iNOS showed complete colocalization in the retina of OIR rats as reported previously [15,35].
In conclusion, we have shown that hypoxia leads to upregulation of nNOS and iNOS in the OIR rat retinas, which can be prevented by treatment with TA. Further studies should be carried out to elucidate the direct responsibility of NO and NOS activity in OIR rats, including identifying the molecular mechanism of TA-induced alterations of NOS systems.


 XML Download
XML Download