

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Proprioceptors exist in extraocular muscles (EOMs) in mammals, though there are differences in kinds and numbers between species. It is believed that they detect small changes of length and tension of EOMs. Moreover, the signals sent to the brain are used to provide information about the fine movement and position of the eye.1 Many reports strongly suggested that the ophthalmic branch of the trigeminal nerve carries proprioceptive afferents.2
There are three types of proprioceptors in human EOM: muscle spindle, the Golgi tendon organ (GTO), and palisade endings.2 The number and location of muscle spindles is variable according to the kinds of EOM.3 Most investigators have asserted that GTO does not exist in human EOM.4,5
Feline and human palisade endings are sensory terminals that are found richly at the myotendinous junction of each EOM and which may be disturbed at the time of strabismus surgery as previously described.5-7 Steinbach and Smith8 have shown that the ability to recognize their eye position is altered in patients who have undergone repeated extraocular surgery. Steinbach et al5 reported that marginal myotomy is more proprioceptively "de-afferenting" than the recession. Most previous papers on myotendinous nerve endings have focused on only normal morphological characteristics or variation according to species. To the best of our knowledge, the morphological changes of nerve endings at myotendinous junction after strabismus surgery have not been investigated. The purpose of this study was to verify the postoperative ultrastructural changes of the numerous myotendinous nerve endings of EOMs according to the recession, and the resection procedures which are most commonly performed. Cats were chosen as the study subjects because feline palisade endings have similar morphological features to those in humans in terms of their traveling course.4
MATERIALS AND METHODS
Eight eyes of four cats (eight lateral recti and eight medial recti) were used and divided into three groups. Under general anesthesia with an intramuscular injection of a 1:1 mixture of ketamine HCl (40 mg/kg) and diazepam (0.05 mg/kg), eight lateral recti were recessed in group A (recession group), four medial recti were resected by 10 mm from insertion to include the myotendinous junction in group B (conventional resection group), and four medial recti were resected by 4 mm of muscle bellies only 10 mm from insertion, without disturbing the myotendinous junction in group C (modified resection group) (Fig. 1). To compare these three groups, the initially resected muscle tissues from group B were used as control.
After surgery, a subconjunctival injection of gentamicin was given and 0.5% ofloxacin eyedrops were instilled into the operated eye two times a day for 3 days.
Four weeks after surgery, the cats were sacrificed by an overdose of pentothal sodium. Each proximal and distal section 2 mm from the myotendinous junction of the operated muscles in groups A and C was cut for specimen preparation. However in group B, because the myotendinous junction might be removed, 4 mm from the insertional area was cut for specimen preparation. Particular care was taken to preserve both the proximal and distal ends of the specimens. All specimens were gently stretched with two forceps during the fixation process to ensure natural anatomic relationships. Proceeding from proximal to distal ends of the specimen, each block was scanned for the presence of nerve fibers, and a step series of semithin cross sections (1 µm) was stained with toluidine blue for light microscopic analysis.
Light Microscopy: Immediately after surgery, semithin transverse sections of resected muscle tissue from group B were examined and photographed under a light microscope after staining with toluidine blue for gross observation of muscle fibers, the myotendinous junction, and the location of nerve endings. The semithin transverse sections of the other specimens obtained from groups A, B and C at four weeks after surgery were also stained with toluidine blue to locate these nerve endings.
Electron Microscopy: Each block was fixed for 4 hours at 4℃ in a mixture of 2% glutaraldehyde and 2% paraformaldehyde in 0.1M sodium phosphate buffer (pH 7.3). Then, the specimens were postfixed with 1% OsO4 at room temperature in the same buffer for 2 hours, dehydrated in a graded series of ethanol, and embedded in Poly-bed 812. Ultrathin transverse sections were stained with uranyl acetate and lead citrate and observed under electron microscope (Hitachi, H-600).
RESULTS
Eight lateral recti were located about 10-12 mm from the corneal limbus, whereas eight medial recti were located about 2 mm from the corneal limbus and were mixed with a thick and complex membranous structure. It was difficult to find the myotendinous junction due to the thick membranous structure around the muscle fibers. Nevertheless, we got used to finding it by noticing a very narrow and long tendinous area (about 10mm long) and a wide muscular area.
Light Microscopy: A series of normal nerve endings was found in the distal myotendinous junction of the medial rectus obtained from the initially resected tissue from group B (Fig. 2). After four weeks, numerous nerve endings located in the distal myotendinous junctional area were found in groups A and C, whereas in group B, no neural structures were found in any section.
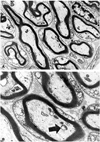
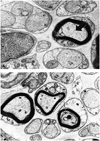
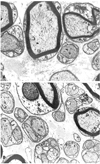
Electron Microscopy: In group A with recession, the overall neural structures were well maintained, but the following slight changes of the axonal cytoskeleton were noted: microtubules were decreased and neurofilaments increased in the axonal cytoplasm, the density of the cytoskeleton was decreased, and overall myelin thickness was regular (Fig. 3). In group B with conventional resection, only muscle fibers were found without any regeneration of neural sprouts. No neural structures were found even under electron microscopy (Fig. 4). In group C with modified resection, many axonal degenerative findings were seen including cytoskeletal changes such as increased neurofilaments and decreased microtubules, and even axonal disintegration was also seen (Fig. 5a). Typical axonal degenerative findings such as axonal shrinkage from myelin was noted (Fig. 5b). In the normal group, the integrity of axon and myelin was well preserved (Fig. 6a, 6b).
DISCUSSION
There are two source of information from which the brain may determine eye position and receive spatial information clues in addition to efferent outflow: visual input from retina and proprioceptive information from EOMs.9 Skavenski et al10 showed that when these two flow signals conflict, the visual input signal is the stronger. Steinbach and Smith8 proposed that proprioceptive information acts as a long-term calibrator that maintains stability and conjugacy of gaze.
Steinbach et al5 found that myotomy of a muscle had a greater effect in de-afferenting proprioception than recession, presumably because of greater destruction of palisade endings by the former procedure. Corsi et al12 reported that the sensorial structures located at the myotendinous junction of EOMs of patients with congenital strabismus had been altered and disorganized, so they recommended that a less invasive procedure such as tendon lengthening or recession might be preferable to other procedures (myotomy, resection) which cause severe structural derangement of the palisade endings.
The kind and number of proprioceptors in mammals are different according to the species. Lukas et al13 noted that GTO was well demonstrated in humans, monkeys, and sheep. However, Steinbach et al5 could not find GTO at the site of strabismus surgery in humans, and instead palisade endings were shown to be relatively abundant at the myotendinous junction, which is the site of many strabismus surgeries.
Richmond et al4 explained that the structure of palisade endings was a single myelinated axon running forward in the tendon, mainly parallel with the longitudinal axis of muscles, turning back to the myotendinous junction, and ramified at this site. Therefore, it appeared as an interwoven network of fine neural filaments, and feline palisade endings had similar morphological features to those in humans in terms of their traveling course, as previously described.4
The location of the myotendinous junction is variable according to rectus muscles and species, but, generally in humans, the tendon length of the medial rectus is the shortest (1-7 mm) and that of the lateral rectus is the longest (4-11 mm).14
Recession is a standard and frequently used technique for weakening a rectus muscle with less muscle damage due to the fact that the muscles are merely moved backwards from insertion. This study revealed that the nerve endings at the myotendinous junction of the lateral rectus in the recession group were well preserved, except for showing a little degeneration in the cytoskeleton. This may have been due to stimulation of the main axon supplying the palisade endings during surgery or minor injury during specimen preparation.
On the other hand, in the conventional resection procedure which is also frequently used, all or part of the tendon, with or without some muscle fiber included, is removed. So we cannot avoid disrupting or even removing the myotendinous junction. It also needs more extensive dissection than recession to augment its effect and reduce the conjunctival lumpiness. Because feline medial recti have thicker membranous structures than lateral recti, we chose the medial recti for resection with extensive dissection.
After severe neural injury, especially with clear transection of the peripheral nerve, the gap separating the two neural aspects is closed by the proliferation of Schwann cells and fibroblasts, which occurs roughly by day 20. On day 100, the neural continuity is established; by day 200, the myelin is established.15 The mass of Schwann cells and a minor amount of cellular debris are formed at 2-4 weeks after neural insult. Degeneration and regeneration of nerves coexist in this period.15 We therefore obtained the specimens at 4 weeks to see active regeneration. However, no neural structure was found in the conventional resection group within 4 mm from insertion. After peripheral nerve transection, as many as 25 sprouts may arise from a single axon.16 Many regenerated axons may reach a termination which is inappropriate and be lost in scar tissue.17 Because the main single axon supplying the palisade endings is located in the margin of each muscle, it might be removed during surgery in dissection, or in specimen preparation. Nevertheless, these results might change with longer follow up and examination of more than 4 mm of the proximal part of the muscle bellies. It was convincing that palisade endings removed during conventional resection would not regenerate into their original form.
A modified resection procedure, as previously described18,19 (rectus resection without scleral sutures), is used only when there is a high risk of scleral perforation. In our study, modified resection was an experimental procedure designed to preserve the myotendinous junction. So 4 mm of muscle bellies proximal to the myotendinous junction was resected. As a result, the nerve ending structure was well preserved grossly, but many axonal degenerative findings were seen. It should be pointed out that the main axon might be cut during this procedure, so axonal degeneration might be progressed anterogradely in nerve endings at the myotendinous junction. Axonal degeneration may result from any pathologic process in which the primary insult is directed toward the axon itself or to the cell body of the axon.20 The ultrastructural features of axonal degeneration include an initial increase in neurofilaments followed by a decrease of axonal organelles, retraction or shrinkage of axon from the myelin sheath, disintegration of the axon, Schwann cell proliferation in distal portions of axons and variable regeneration in the form of axonal sprouting.20-21 Many nerve fibers in the distal myotendon of group C had the above characteristics, which certainly implies axonal degeneration. Sas and Schab22 noted the degeneration of palisade endings after destruction of the 3rd, 4th and 6th nerve nuclei.
We could also expect that large vascular structures such as the muscular branch of the ophthalmic artery or ciliary artery were cut during this experimental procedure, so that vascular infarction resulted distal to the point of resection. It is an object for further study whether modified resection causes possible circulatory disturbance or infarct of muscle. We could not determine whether or not the modified resection can be expected to produce functionally similar results to the conventional resection.
Ultrastructural studies investigating morphological change are not well-suited for quantitative study due to their staining method and teasing technique.23 Furthermore, it is very difficult to examine because there are numerous unspecified structures which can be suspected as nerve fiber. Therefore, we did not count the number of degenerating nerve endings present in each specimen.
We could not exclude the possibility that motor nerves were included in this study because the axon bundles supplying palisade endings are often found mixed with those giving the motor endings,as previously described.23 However, although motor and sensory nerves are indistinguishable ultrastrucuturally,24 it is convincing that there were no differences in the ultrastructural changes of degeneration between the palisade endings and motor nerves. Additionally, motor nerve damage or paralysis would also produce disturbance in trigeminal-oculomotor synkinesis.8,24
We confirmed that nerve endings at the myotendinous junction can be damaged in strabismus surgery, and also that recession is a more effective procedure to preserve nerve endings than resection. Nevertheless, it would be premature to use these results to decide on preferred forms or techniques of strabismus surgery in terms of surgical goals and patient outcomes. Further study is necessary to investigate how these aspects of nerve damage affect clinical manifestations.



 XML Download
XML Download