

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Na+/K+-ATPase (NKA; sodium-potassium pump) is abundantly expressed in the basolateral membrane of epithelial cells and maintains cell homeostasis by transporting three Na+ molecules out and two K+ molecules into a cell, using the energy from the hydrolysis of adenosine triphosphate (ATP) (12). NKA is the primary active transporter that produces the driving force for the activation of other secondary ion transporters on cell membranes (12). The function of NKA controls a variety of ion and metabolite transport systems. Moreover, this pump alone accounts for approximately 30% of ATP consumption in animal cells for the purposes of ion regulation to maintain intracellular homeostasis (34).
NKA is also a potential regulator of tight junction formation and function (567). Tight junctions function as urothelial barriers between urine and the underlying bladder (89). They also play a critical role in the biogenesis of polarized epithelial cells during early vertebrate development and during recovery from tissue damage after ischemic or toxic injury (210). Previous studies have reported that tight junction protein expression is decreased in the bladder urothelium (111213) and that increased potassium leakage causes clinical symptoms in patients with interstitial cystitis/painful bladder syndrome (IC/PBS) (14). Therefore, studies of changes in NKA function in bladder tissues may be important for development of appropriate therapeutic strategies in patients with bladder disease.
Here, we aimed to investigate changes in NKA expression and activity in bladder biopsies from patients with IC/PBS. To the best of our knowledge, this is the first study to investigate changes in NKA function in patients with this bladder disease.
MATERIALS AND METHODS
Patients and tissue samples
Bladder specimens were obtained from 85 patients (78 women and 7 men) with IC/PBS who were undergoing cystoscopy under anesthesia for diagnosis of IC/PBS or for therapeutic bladder hydrodistention. All IC/PBS patients were defined with the cystoscopic criteria of National Institute of Diabetes, Digestive, and Kidney Diseases (15) and had experienced clinical symptoms for more than 6 months. The average age of patients in the cohort was 41.2 ± 9.4 years. Patient backgrounds, including clinical symptoms and past therapeutic history, are shown in Table 1. All patients underwent medical history collection, physical examination, urinalysis, and culture for exclusion screening and were required to stop the use of intravesical instillation drugs for at least 1 month before biopsy. IC/PBS symptoms were evaluated by questionnaires assessing O'Leary-Sant symptoms, problem indexes, and pelvic pain and urgency/frequency (PUF) (16). In the study group, a cold-cup biopsy was taken at sites demonstrating glomerulation after hydrodistension. Control specimens were obtained from 20 patients (median age: 42.7 years, range: 38–58 years; 18 women and 2 men who had undergone transurethral resection of the prostate more than 1 year prior) undergoing bladder neck suspension for stress urinary incontinence who had no symptoms of IC/PBS and showed no evidence of bladder mucosal disease after hydrodistension under cystoscopy. Hydrodistension was performed using normal saline for infusion fluid under gravity (100 cm pressure) until complete. The distension time was started, and metal compression was applied to prevent leakage for 2 minutes (17). All tissues were stored at -80°C for immunoblotting and NKA activity assays or fixed in 10% formalin for immunostaining. All specimens were collected after obtaining informed written consent from patients.
Table 1
Demographic and clinicopathological data for patients with interstitial cystitis/painful bladder syndrome

![]()
Antibodies
Four primary antibodies were used in this study as follows. Mouse monoclonal anti-NKA antibodies (α5; Developmental Studies Hybridoma Bank, Iowa City, IA, USA) were used for immunoblotting (dilution 1:200) and immunohistochemical (IHC) staining (dilution 1:200). Rabbit polyclonal anti-NKA antibodies (EP1845Y; Abcam, Cambridge, MA, USA) were used for immunofluorescence (IF) staining (dilution 1:50) to detect all α-isoforms of NKA. Additionally, mouse monoclonal anti-cytokeratin 7 (CK7) antibodies (NCL-L-CK7-OVTL; Novocastra, Milton Keynes, UK) was used for IF staining (dilution 1:50) to identify bladder uroepithelium (1819). Rabbit polyclonal anti-actin antibodies (sc-1616-R; Santa Cruz Biotechnology, Santa Cruz, CA, USA) raised against the C-terminus of human ACTB were used as a loading control for immunoblotting (dilution 1:5,000).
The secondary antibodies for immunoblotting were horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG or goat anti-rabbit IgG (#0031430 or #0031460, respectively; Pierce, Hercules, CA, USA). The secondary antibodies used for immunostaining included a commercial kit (PicTure; Zymed, South San Francisco, CA, USA) for IHC staining and Alexa Fluor 488-conjugated goat anti-mouse or Alexa Fluor 546-conjugated goat anti-rabbit antibodies (Molecular Probes, Eugene, OR, USA; dilution 1:50 and 1:200, respectively) for IF staining.
Preparation of tissue homogenates
Each sample was mixed with 200 µL SEID buffer (300 mM sucrose, 20 mM EDTA, 100 mM imidazole, 0.1% sodium deoxycholate, pH 7.5) containing proteinase inhibitor (#11836145001; Roche, Indianapolis, IN, USA; v/v: 25/1) and then homogenized on ice. The homogenate was centrifuged at 5,000 × g at 4°C for 5 minutes. The supernatants were collected and used for immunoblotting, NKA activity assays, or protein concentration analysis. Protein concentrations were determined using a BCA protein assay kit (#23225; Pierce) with bovine serum albumin (#23209; Pierce) as a standard.
Immunoblotting
The method for immunoblotting was modified from our previous studies (1820). In brief, aliquots containing 50 µg of homogenates prepared as stated above were added to the sample buffer and heated to 60°C for 15 minutes. After electrophoresis and transformation, the blots were incubated sequentially in blocking buffer, primary antibodies (α5 or actin), and secondary antibodies. Finally, protein bands were detected using a SuperSignal West Pico Detection Kit (#34082; Pierce) and imaged using a cooling-CCD image sensor (ChemiDoc XRS+; Bio-Rad, Hercules, CA, USA) with specialized software (Quantity One version 4.6.8; Bio-Rad). Immunoreactive bands were analyzed using Image Lab software version 3.0 (Bio-Rad). The results were converted to numerical values in order to compare the relative protein abundance of the immunoreactive bands.
NKA activity assay
Enzyme activity was measured using the NADH-linked method. ADP derived from the hydrolysis of ATP by ATPase was enzymatically coupled to the oxidation of reduced NADH using lactate dehydrogenase (LDH) and pyruvate kinase (PK). In this study, the method for assaying NKA activity was modified from previous studies (2122). The assay reaction was started by adding 10 µL of sample supernatant and 200 µL of reaction mixture (0.38 mM ATP, 1.50 mM phosphoenolpyruvate, 0.24 mM NADH, 2.48 U/mL LDH, 2.70 U/mL: PK, 47.3 mM NaCl, 2.63 mM MgCl2·6H2O, 10.5 mM KCl, 50 mM imidazole, pH 7.5) with or without 0.75 mM of ouabain (NKA inhibitor) to each well. The samples were examined every 15 seconds for up to 20 minutes using a VERSAmax microplate reader (Molecular Devices, Sunnyvale, CA, USA) at 340 nm and 37°C. The linear rate from 5 to 20 minutes for each pair of triplicate wells was determined. NKA activity was calculated as the difference in the slope of ATP hydrolysis (NADH reduction) in the presence or absence of ouabain, with activity expressed as µmol ADP per µg protein per hour.
IHC staining
The staining and microscopy methods were modified from our previous studies (1923). The formalin-fixed, paraffin-embedded sections (4 µm) of biopsy samples were dewaxed and then rinsed with phosphate-buffered saline. Endogenous peroxidase was inactivated by incubating the sections with 3% hydrogen peroxide. The sections were stained with anti-NKA (α5) or phosphate-buffered saline (as a negative control) before analysis using a commercial kit. Sections were observed by optic microscopy (BX50; Olympus, Tokyo, Japan) with cooling CCD (DP72; Olympus) using CellSens software (CellSens standard version 1.4; Olympus).
Multiple IF staining
The method for IF staining was modified from our previous studies (1820). In brief, paraffin-embedded sections were incubated at 4°C overnight with diluted primary antibodies (α5 or CK7) and then exposed to the appropriate secondary antibodies for 1 h. Finally, the sections were covered with a coverslip using mounting solution (DAPI Fluoromount G, #0100-20; SouthernBiotech, Birmingham, AL, USA). To identify and compare the localization of specific proteins, IF-stained samples were observed with a fluorescent microscope (BX50; Olympus) with cooling CCD (DP72; Olympus) using CellSens software (CellSens standard version 1.4; Olympus). Micrographs were subsequently merged to visualize differently colored labels simultaneously.
Statistical analysis
Differences between the groups were analyzed using Mann-Whitney U tests, and all values are expressed as the mean ± standard error (SE). Differences with P values of less than 0.05 were considered significant. The analyses were performed with SPSS 12.0 software (SPSS, Chicago, IL, USA).
RESULTS
The protein levels (1.08 ± 0.06 vs. 2.39 ± 0.29, respectively) and activity (0.60 ± 0.04 vs. 1.81 ± 0.18 µmol ADP/mg protein/hour, respectively) of NKA were significantly lower in the study group (about 45% and 1/3-fold, respectively) than in the control group (P < 0.05; Fig. 1 and 2). Additionally, the bladder urothelium was thinner in patients with IC/PBS (Fig. 3A) compared with patients in the control group (Fig. 3C), and IHC staining for NKA (red) showed weaker immunoreactivity in the study group (Fig. 3B) than in the control group (Fig. 3D). Furthermore, multiple IF staining for NKA (green) and CK7 (uroepithelial marker, red) demonstrated that these two targets colocalized in the bladder urothelium of patients with IC/PBS (Fig. 4). These staining results revealed that NKA was predominantly expressed in the bladder urothelium (Fig. 3 and 4). Taken together, these data showed that the expression and activity of NKA were lower in bladders of patients with IC/PBS than in control individuals.
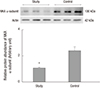 | Fig. 1Representative immunoblot and relative intensities of the Na+/K+-ATPase (NKA) α-subunit in patients in the study and control groups. Actin was used as the loading control. Values are the mean ± SE. The asterisk indicates a significant difference compared with the control group.
|
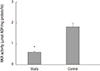 | Fig. 2Na+/K+-ATPase (NKA) activity in the study and control groups. Values are the mean ± SE. The asterisk indicates a significant difference compared with the control group.
|
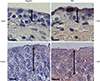
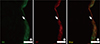
DISCUSSION
NKA is expressed in the basolateral plasma membrane of most epithelial cells (12) and plays a key role in maintaining cell homeostasis in the epithelium of organs and tissues, including the bladder (25). In the present study, our results showed that NKA was predominantly expressed in the bladder urothelium of both the study and control groups, similar to the results of other mammalian studies (242526). Moreover, the expression and enzyme activity of NKA were decreased in patients with IC/PBS compared with those in the control group. We found no evidence that this pump compensated for potassium leakage in the study group. Thus, these results implied that bladder urothelial NKA was dysfunctional in patients with IC/PBS.
Rajasekaran et al. reported that NKA activity is necessary for tight junction formation in epithelial cells (57). The downregulation of tight junction proteins, such as zonula occludens-1, occludin, and E-cadherin, in the bladder urothelium in patients with IC/PBS has been reported previously (1927). Moreover, inhibition of NKA ion transport function increases the permeability of tight junctions to ionic and nonionic solutes (2829). The increased permeability and decreased tight junction formation in bladder urothelial cells was confirmed in patients with IC/PBS (11) and was shown to permit the migration of urinary solutes (81214). In particular, high potassium levels in urine (i.e., 24–133 mEq/L) (814) could easily lead to leakage into interstitial tissue through dysfunctional bladder urothelium, causing depolarization of nerves and detrusor contractions and resulting in bladder pain, urgency, and increased frequency of urination in patient with IC/PBS (303132).
Previous studies have reported that NKA is a signaling molecule involved in multiple cell signaling pathways. Inhibition of NKA function activates certain signaling, including Ras, phosphoinositol 3 kinase, mitogen-activated protein kinase (MAPK) and apoptotic pathway (733). Higher levels of bladder urothelial apoptosis have been observed in patients with IC/PBS (1827). Therefore, dysfunction of bladder urothelial NKA may contribute to the observed increase in apoptosis.
In addition, NKA requires direct coupling with metabolic energy (i.e., ATP) (12). Our previous study showed that hypoxia-inducible factor-1α expression is increased in the bladder tissues of patients with IC/PBS (20), indicating that hypoxia occurring in these tissues may cause lower energy (ATP) production and impair NKA function (343536). Taken together, these data suggested that decreased NKA expression and activity may be associated with tissues hypoxia (2023), which could decrease tight junction formation (19) and increase apoptosis (18). These events would result in the formation of thinner, dysfunctional bladder urothelium in patients with IC/PBS.
There were several potential limitations to this study. First, patients' clinical symptoms and disease duration may have been subjected to selection bias. Second, there are still no uniform standard criteria and definitions with which to diagnose this disease. Third, the proposed relationships among hypoxia, apoptosis, tight junctions, and NKA are based on the results of our previous studies (18192023).
In conclusion, we found that the expression and activity of NKA were decreased in bladder biopsies from patients with IC/PBS and that NKA was predominantly localized on the bladder urothelium. These findings suggest that impaired NKA function in the bladder urothelium may contribute to the diseased state in patients with IC/PBS.
 XML Download
XML Download