

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Parkinson’s disease (PD) is a serious chronic neurodegenerative disease that is caused by the loss of dopaminergic neurons in the substantia nigra. A depletion of dopamine in the striatum leads to parkinsonism (1). Different drug treatments have been proposed, but most chemicals have shown limited efficacy (2). In order to control the emergence of PD, cell therapy has been suggested as a promising alternative treatment. Data from animal models and clinical trials have suggested that cell replacement therapies may be an effective treatment for this disorder (34). However, cell replacement therapy needs to consider immune rejection, teratoma formation or ethical issues (56).
Autologous mesenchymal stem cells (MSCs) were frequently tried as the reliable cell source avoiding above limitations for PD treatment (7). MSCs are multipotent stem cells that can be differentiated into neurons. In order to obtain autologous MSCs, invasive operations are usually required. However, to establish a standard and applicably safe cell therapy, noninvasive operations are ideal approaches for patients. Human dental pulp stem cells (hDPSCs) have been demonstrated to be a good source for autologous stem cell. The hDPSCs can be obtained noninvasively from deciduous teeth, and maintain their self-renewal capability and multipotency (8910) including neuron-like cells (911). hDPSCs can also be derived from cryopreserved dental pulp (12).
In the present study, we identified the potential of hDPSCs to differentiate into dopaminergic neurons under appropriate conditions in vitro. These results could provide fundamental information regarding the potential of hDPSCs as an alternative stem cell source for PD treatment.
MATERIALS AND METHODS
hDPSC culture
hDPSCs were kindly provided by Prof. Eui Kyun Park, Kyungpook National University, Korea. The hDPSCs were maintained in α-minimum essential medium (α-MEM; Invitrogen Corp., Carlsbad, CA, USA) supplemented with 10% fetal bovine serum (FBS; Invitrogen), 100 units/mL penicillin, and 100 mg/mL streptomycin (HyClone Labs, Thermo Scientific, Rockford, IL, USA). The cells were maintained at 37°C in a humidified 5% CO2 incubator. Cells that had undergone less than five passages were used for this study.
hDPSCs’ differentiation into dopaminergic neurons
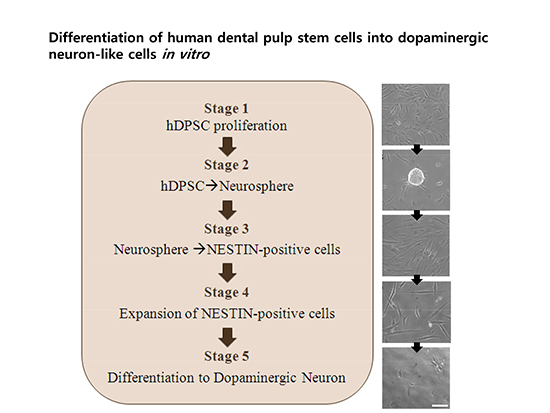
For differentiation into dopaminergic neurons, hDPSCs were treated with the Human/Mouse Dopaminergic Neuron Differentiation Kit (R&D Systems, Abingdon, UK). At stage 1, hDPSCs were preinduced on gelatin-coated plates with recombinant mouse leukemia inhibitory factor (LIF)-containing knockout-ES (KO-ES/LIF) medium, made up of knockout Dulbecco’s modified Eagle’s medium (DMEM), FBS, MEM non-essential AA solution, penicillin-streptomycin-glutamine, and β-mercaptoethanol, for 3–4 days. At stage 2, 2 × 106 cells on 10-cm culture plates were treated with KO-ES medium without LIF for 4 days to induce neurosphere formation. At stage 3, to select for Nestin-positive cells, the neurosphere was settled to the plate with KO-ES medium. After neurosphere attachment to the plate, the medium was replaced with human insulin-transferrin-sodium (ITS)/fibronectin medium containing DMEM/F-12, glucose, l-glutamine, NaHCO3, ITS supplement, and bovine fibronectin for 6–8 days. At stage 4, for Nestin-positive cell expansion, 5 × 105 cells/well were transferred into poly-l-ornithine/fibronectin-coated 24-well plates with N-2-supplemented (insulin, transferrin, selenite, putrescine, and progesterone) medium that also contained basic fibroblast growth factor (bFGF), FGF-8b, sonic hedgehog-N (Shh-N), and ascorbic acid, for 4–6 days. At stage 5, to differentiate cells into dopaminergic neurons, the cells were cultured in N-2/ascorbic acid medium (without growth factors) for 10–15 days.
Characterization of differentiated hDPSCs by immunocytochemistry
During the five differentiation stages, the neural characteristics of the cells were analyzed on the protein level. The hDPSCs were cultured on a chamber slide for 1 day. Subsequently, the cells were fixed with 4% paraformaldehyde in 0.1 M phosphate buffer (pH = 7.4) and then treated with 4% bovine serum albumin (BSA; Sigma-Aldrich, St. Louis, MO, USA). The cells were then permeabilized with 0.5% Triton-X 100 (Sigma-Aldrich) in phosphate-buffered saline (PBS) for 1 hour. The antibodies were listed under Fig. 1A. The DNA-specific fluorescent dye 4,6-diamidino-2-phenylindole (DAPI; Santa Cruz Biotechnology, Santa Cruz, CA, USA) was used as the counterstain to detect cell nuclei. The substantia nigra of ICR mouse brain tissue was used as the positive control (Fig. 1B).
 | Fig. 1Morphological and immunocytochemical analyses of human dental pulp stem cells (hDPSCs) throughout the various differentiation stages. (A) The neuronal characteristics of hDPSCs were verified at the protein level by immunocytochemical analysis. (B) Substantia nigra of ICR mouse brain tissue was used as the positive control for immunohistochemistry. SSEA4, stage-specific embryonic antigen 4; TuJ1, β-tubulin III; GFAP, glial fibrillary acidic protein; MBP, myelin basic protein; TH, tyrosine hydroxylase; H&E, hematoxylin & eosin.
|
Characterization of differentiated hDPSCs by flow cytometry
The cultured hDPSCs in neural inductive condition were dissociated with accutase (Millipore, Billerica, MA, USA). Acquired single cells were collected, resuspended with 2% FBS in PBS, and stained with a surface antigen, SSEA4. In order to stain for intracellular antigens (Nestin, TuJ1, GFAP, MBP, and TH), cells were fixed with 4% paraformaldehyde in 0.1 M phosphate buffer, washed twice, and then permeabilized with 0.2% Triton X-100 in PBS on ice. Approximately 5 × 105 cells were collected and divided into 1.5-mL tubes, into which the same antibodies as those used for immunocytochemistry were added at 1:40 dilutions for the flow cytometry analysis. Stained hDPSCs were analyzed for their neural characteristics on a fluorescence-activated cell sorter calibur instrument (BD Biosciences, San Jose, CA, USA).
Characterization of differentiated hDPSCs by real-time PCR
At the transcript level, the neural characteristics were analyzed by real-time PCR. Total RNA was extracted from cultured cells by using the TRIzol Reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. One microgram of RNA was reverse-transcribed with a Maxime RT Premix Kit (iNtRON Biotechnology, Korea), using a C1000 thermal cycler (Bio-Rad Laboratories, Hercules, CA, USA). For the real-time PCR analysis, the SYBR Green PCR Master Mix and ABI Prism Sequence Detection System 7000 (PE Applied Biosystems, Foster City, CA, USA) were used. The sequences of the primers (Bioneer, Korea) are shown in Table 1. The PCR cycling conditions were 50°C for 2 minutes, 95°C for 10 minutes, and 40 cycles of 95°C for 15 seconds, 60°C for 1 minute, 95°C for 15 seconds, 60°C for 20 seconds, and 95°C for 15 seconds.
Table 1
Nucleotide sequences of the primers used for real-time PCR

![]()
Tyrosine hydroxylase quantification by enzyme-linked immunosorbent assay
To confirm quantitatively the dopaminergic differentiated hDPSCs, the tyrosine hydroxylase enzyme-link immunosorbent assay (TH ELISA) was performed. Total protein extracted from cells, undifferentiated hDPSCs (stage 1), and fully differentiated hDPSCs (stages 5) were prepared using RIPA lysis buffer (GenDEPOT Laboratories, Barker, TX, USA). Protein concentrations were determined by the Bradford method (Sigma-Aldrich), using BSA as the standard (Thermo Scientific). After verifying the final protein concentrations, the same concentration of protein was used in all the samples. All samples were used to measure the levels of TH, according to the manufacturer’s protocol (USCNK Life Science Inc., Wuhan, China).
RESULTS
Morphological analysis of differentiating hDPSCs
Following the addition of inducing agents into the medium, the morphology of differentiated hDPSCs changed as dopaminergic neurons in vitro through the five differentiation stages (Fig. 1A). At preinduction stage 1, the hDPSCs showed a flat and cobble-stone-like morphology. Neurospheres were formed at stage 2. Then, the monolayer cells were spread out from the attached neurospheres, showing a fibroblastic morphology at stage 3. The expanded cells showed a neural progenitor shape, such as spindle-shaped cells with bipolar processes or pyramidal cells, at stage 4. Finally, the cells were transformed into a dopaminergic-like neuron shape with a condensed and retracted cell body at stage 5, and adopted a radial glial-like appearance.
Immunocytochemical analysis of hDPSC differentiation
SSEA4 (stem cell marker) was expressed from stages 1 to 4, and the SSEA4-positive cell number decreased gradually and the expression was not detected at stage 5. In the case of NESTIN (neural stem cell marker) and TUJ1 (early neural marker), they were expressed in various cell numbers compared with the other antibody groups showing fibroblast-like morphology, and the expression continued through all the five stages. A small portion of hDPSCs expressed GFAP (mature neural markers for astrocyte) before and after neuronal differentiation, while MBP (mature neural markers for oligodendrocyte)-positive cells appeared abundantly from stage 1 onward, and cells positive for both these antibodies appeared only round shaped. TH (dopaminergic marker)-positive cells were appeared at stage 5 only (Fig. 1A). As the positive control for several antibodies, the substantia nigra of ICR mouse brain tissue was used (Fig. 1B).
Flow cytometric analysis of hDPSC differentiation
hDPSCs were differentiated into neural progenitor, early neuron, and dopaminergic cells in culture. The results showed that the percentage of SSEA4 through the stages decreased gradually, and the value at stage 5 was 1.76%, meaning that hDPSCs were losing their stem cell character. NESTIN- and TUJ1-positive cells at stage 1 accounted for more than 90% of the hDPSCs, but the value decreased significantly when cells started forming the neurosphere; however, from stage 3 onward, their expression levels increased gradually and the population reached 100% at stage 5. GFAP-positive cells made up 6.26% at stage 1 and the population did not show a significant change in number until stage 5. The initial MBP-positive cell population was 32.88% and reached 94.13% at the final stage. In the case of TH-positive cells, the initial expression level was very low (1.02%), but the population increased significantly during the experiment, giving a final value of 14.49% (Table 2). The relative expression percentage of each value was compared with mouse IgG1 isotype as the control.
Table 2
Flow cytometric results of target genes through differentiational stages of hDPSCs

![]()
Genotypic analyses of hDPSCs by real-time PCR
The expression of Vimentin (neural stem cell marker) was constant, whereas the expressions of NESTIN and PAX6 (neural lineage marker) increased gradually over the course of time. TUJ1, MAP2, GFAP, MBP, O4 (oligodendrocyte marker), and TH showed maximized expressions at stage 4 (Fig. 2).

Identification of differentiated hDPSCs by TH ELISA
The quantification of TH from cell lysates was evaluated with an ELISA kit (Fig. 3). As compared with stage 1 (0.110 ± 0.032 ng/mL), the TH value at stage 5 increased to 0.526 ± 0.033 ng/mL. The P value for the two-tailed test was less than 0.0001. These results suggest that hDPSC-derived dopaminergic neurons possess the ability to synthesize dopamine.

DISCUSSION
Currently, hDPSCs have been shown to be able to differentiate into neural precursors or neurotrophic support for dopaminergic neurons, but not into mature dopaminergic neurons (13). In the present study, we have provided novel evidence that dopaminergic neurons and several neuronal cells can be generated from hDPSCs by in vitro stimulation. To explore the differentiation of hDPSCs, specific markers of the various neuron developmental stages were selected; namely, SSEA4 (a marker for mesenchymal stem cells), Nestin (a neural stem cell and progenitor cell marker), Pax6 (a neuronal lineage marker), TUJ1 (an early marker for neurons), MAP2 (a marker for mature neurons), GFAP (a mature neuron marker for astrocytes), MBP and O4 (mature neuron markers for oligodendrocytes), and TH (an enzyme required for the generation of dopamine and thus considered as a marker for dopaminergic neurons) (1415).
Protein level analysis of the differentiating hDPSCs was verified by immunocytochemical analysis. The type and level of protein expression for neural-specific markers depend on the various differentiation stages. Expression of the stem cell maker was observed at the early stages, but the frequency decreased gradually. Expressions of the early neural stem cell marker and early neural marker remained steady throughout the various stages. The mature neural markers were highly expressed from stage 3 onward. Most importantly, expression of the dopaminergic neuron marker was found at stage 5. To confirm the immunocytochemistry results, we performed a flow cytometry analysis. Quantitative cell count analyses indicated that of the treated hDPSCs, 100% expressed Nestin and TUJ1, 8.24% expressed GFAP, and 94.13% expressed MBP, and of this population, 14.49% expressed TH. This result was consistent with the immunocytochemistry experiments.
Real-time PCR analysis indicated that when hDPSCs were cultured in neuronal inductive medium, the Nestin transcript levels appeared to be upregulated from stage 3 onward, whereas there was only a minor alteration in the expression of VIMENTIN. The PAX6, TuJ1, MAP2, O4, MBP, and TH transcript levels were all increased in the differentiation medium, which indicated that hDPSC-derived spheres could generate mature neurons and a number of committed differentiated dopaminergic neurons. Interestingly, the induced hDPSCs expressed high levels of MBP and O4 transcripts, but low levels of GFAP, which is consistent with the ability of the induction cocktail to trans-differentiate the hDPSCs into an oligodendrocyte lineage. At stage 5, GFAP expression had decreased to about 8%, meaning that the astrocyte population was relatively low.
To determine whether the induced hDPSCs release TH, we measured the intracellular levels of TH at stages 1 and 5, using an ELISA kit. The TH level was significantly enhanced at stage 5, increasing to 0.526 ± 0.033 ng/mL from 0.110 ± 0.032 ng/mL at stage 1. This indicated that hDPSCs could potentially differentiate into dopaminergic neuronal-like cells in an appropriate environment. This 5-step method produced about 1.02% TH-positive neurons at an efficiency of about 92.55% of TUJ1-positive neurons in the presence of ITS in the medium. Moreover, when FGF basic, FGF-8b, Shh-N, and ascorbic acid were added to the medium, the production increased to about 14.5% TH-positive neurons at an efficiency of 100% of TUJ1-positive neurons. TH was expressed in hDPSCs even in the absence of the differentiation factors retinoic acid and brain-derived neurotrophic factor (16).
Signaling with trophic factors plays an important role in the differentiation of stem cells (16). During the procedure, we simulated the induction and differentiation of hDPSCs using several growth factors. For the preinduction stage 1 of hDPSCs, medium containing β-mercaptoethanol, serum, and LIF was used (17). β-Mercaptoethanol and serum are survival factors that support the viability and differentiation of neurons (1819). LIF is required for appropriate changes in neuronal gene expression to ensure the survival and maintenance of neurons (20), as well as to promote neural stem cell self-renewal, preventing the emergence of more differentiated cell types (21). hDPSCs formed the neurosphere in the KO-ES medium without LIF/retinoic acid (RA) at stage 2. LIF impairs neurosphere formation in vitro, even at a very low dose (21). Moreover, the hDPSCs showed neurosphere formation without RA treatment. The avoidance of RA treatment is based on the fact that RA is a negative factor for dopaminergic neuron generation (22). The neurosphere is a nonadhesive aggregate that is composed of free-floating clusters of neural precursor cells of varying cell types (heterogeneous), including neurons, astrocytes, and oligodendrocytes (23). The neurosphere-like aggregates of stem cells are an important property of neural stem cells (24). This formation of floating spheres under the appropriate medium means that hDPSCs might possess the potential to differentiate into neural stem cells. In the neurosphere stage, the SSEA4, Nestin, TuJ1, and GFAP expression levels were decreased, while those of MBP and TH were increased, meaning that the neurosphere niche is different to that of the stage 1 environment.
After the neurosphere had attached to the plate, neural progenitor cells spread out into a monolayer. At stage 3, ITS and N-2 were used to select and enrich for neural stem cell populations. The included fibronectin provides support for the cell attachment and spreading. The FGF and Shh-N contained in the medium induce effective neuronal differentiation because without these factors, neuronal morphology is not observed (25). In the late stages, hDPSC-derived spheres were induced into dopaminergic neurons with a cocktail of Shh-N, FGF-8b, FGF basic, and ascorbic acid. These are thought to work in unison to mediate the induction of dopaminergic cells (26). Under this condition, the hDPSC-derived spheres generated a number of TuJ1- and MAP2-positive neurons, and at the final step, some of them were TH-positive cells, suggesting that the controlled differentiation of hDPSCs by the growth factor cocktail had led to the activation of active TH and the induction of functional neurons (24). Wang et al. (24) showed that TH was expressed only in neurospheres induced by a cocktail of these growth factors.
In this study, we have identified the potential dopaminergic differentiation of hDPSCs, which was confirmed by morphological, immunocytochemical, flow cytometric, real-time PCR, and ELISA analyses. The induced TH-positive neurons were shown of TH. These results suggest that hDPSC-derived dopaminergic neurons possess the ability to synthesize dopamine. Most of the dopaminergic differentiation studies have used ES cells up to now. However, our study suggests that hDPSCs can be a promising autologous cell source for the treatment of PD.
For further study, it will be important to determine whether these cells possess the electrophysiological characteristics of neurons, as well as to investigate the therapeutic efficacy of hDPSCs in alleviating PD in an animal model through transplantation.
 XML Download
XML Download