

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Pancreatic cancer is one of the leading causes of cancer death, with a 5-year survival rate of < 5% (12). Less than 20% of patients present with potentially resectable pancreatic cancer, whereas the remaining patients have locally advanced, unresectable, or metastatic disease (3). The disease is characterized by rapid tumor spread and dismal prognosis. Moreover, most patients are diagnosed at an advanced stage and are not candidates for local treatments, such as surgical resection and irradiation (4). Therefore, identifying additional target molecules that are crucially involved in the growth of pancreatic cancer cell growth is necessary in developing an effective treatment.
Plenty of eukaryotic initiation factors (eIFs) are involved in translation initiation, which is the significant rate-limiting step in the process of protein translation (5). Among all eIFs, eIF3 is the largest family member and contains at least 13 subunits, consecutively designated eIF3a to eIF3m (67). The eIF3 family members are demonstrated to play two important roles in protein translation. On one hand, eIF3 can bind onto the 40S ribosome and promote loading of the ternary complex of Met-tRNA/eIF2-GTP, finally forming the 43S preinitiation complex. On the other hand, eIF3 notably assists eIF4 to recruit mRNAs to the 43S complex (8910). The above observations suggest that eIF3 is one of the most important initiation factors in eukaryote.
Among the 13 subunits of eIF3, eIF3a is the largest one and also known as p170 (11). eIF3a is critical for ribosomal subunit joining and recruiting mRNA to the ribosome, as well as for the regulation of various gene products, in which ribonucleotide reductase M2 and some DNA repair molecules are involved (121314), the latter of which suggests that overexpression of eIF3a may suppress cellular protection against DNA damages via inhibiting DNA repair, leading to higher frequency of gene mutation for tumorigenesis and lower sensitivity of cancer cells to DNA-damaging-associated anticancer drugs (1516). Thus, eIF3a expression is associated with tumorigenesis (13), metastasis (11), cancer prognosis (17) and malignant transformation (18). Shen et al. (19) demonstrated that eIF3a functioned as a regulator of xeroderma pigmentosum complementation group C (XPC) and p27Kip1 translation in ovarian cancer. eIF3a was also increased in human urinary bladder cancer and affected its phenotype independent of translation initiation (20). However, the detailed role of eIF3a in other cancers such as pancreatic cancer remains largely unknown.
Herein, eIF3a expression level in pancreatic cancer tissues and seven pancreatic cancer cell lines were examined in this study. Furthermore, the effects of eIF3a knockdown on tumorigenic ability and cell migration and invasion were assessed in pancreatic cancer cells. The tumorigenic abilities of pancreatic cancer cell lines were also examined after eIF3a knockdown in a nude mouse model. This report represented the first study as far as we know to investigate the role of eIF3a in the cell proliferation and metastasis in pancreatic cancer.
MATERIALS AND METHODS
Cell culture
PANC-1, AsPC-1, SW1990, Capan-1, Miapaca-2, BxPC-3 and L3.6pL cells were purchased from Shanghai Institutes for Biological Sciences (Shanghai, China). Miapaca-2 cells were maintained in RPMI 1640 medium (Gibco, MD, USA). PANC-1, AsPC-1, SW1990, Capan-1, BxPC-3 and L3.6pL were maintained in Dulbecco’s modified Eagle medium (DMEM) (Gibco). All media were supplemented with 10% fetal bovine serum (Gibco), and the cells were cultured in 5% CO2 at 37°C.
Patients and tissue samples
Slides from 140 clinical patients suffering from pancreatic ductal adenocarcinoma were collected (5 slides for each case). Clinicopathological information of these patients were also obtained from the First Affiliated Hospital, Zhejiang University. Another 30 patients who underwent surgery at the First Affiliated Hospital, Zhejiang University from 2010 to 2015 were chosen after histological verification of pancreatic ductal adenocarcinoma. For these 30 patients, fresh cancerous tissues and the paired adjacent non-cancerous tissues were synchronously collected for each patient. None of the 30 patients received preoperative chemotherapy or radiotherapy.
Immunohistochemical analysis and evaluation of eIF3a expression
Immunohistochemical staining was performed using a standard immunoperoxidase staining procedure, and the eIF3a expression in benign and malignant specimens was evaluated as follows: immunoreaction extent, 0 score (positive cells in less than 5% of total); 1 score (positive cells with 5%–25%); 2 scores (positive cells with 25%–50%); and 3 scores (positive cells with greater than 50%); staining intensity, 0 score (no coloration); 1 score (pale yellow); 2 scores (yellow); and 3 scores (claybank). The two scores were then multiplied: 0 score as negative; 1–3 scores as weak; 4–6 scores as moderate; > 6 scores as strong.
RNA isolation and semi-quantitative reverse transcription–PCR
RNAs were isolated from the 30 cases of pancreatic ductal adenocarcinoma. Total RNAs from PANC-1, AsPC-1, SW1990, Capan-1, Miapaca-2, BxPC-3 and L3.6pL cells were extracted using TRIzol solution (Invitrogen, Carlsbad, CA, USA). The primers for eIF3a and β-actin were listed as follows: eIF3a: 5'-ACAGGCAGTGTTTGGAC-3' (forward) and 5'-GAGAATAGCCCGTGAATA-3' (reverse); β-actin: 5'-TGGCACCCAGCACAATGAA-3' (forward) and 5'-CTAAGTCATAGTCCGCCTAGAAGCA-3' (reverse). The PCR was performed as previously described (18). The house-keeping gene β-actin was included as an internal standard.
Western blot analysis
Cells were collected and solubilized with Cell Lysis/Extraction Reagent (Sigma) and with complete protein inhibitor cocktail (Roche, IN, USA). The equal amount of sample was loaded in each well of a 12% gel and subjected to SDS-PAGE. Gels were transferred to polyvinylidene difluoride (PVDF) membrane (Millipore, MA, USA). The membrane was then incubated with primary antibodies against eIF3a (1:1,000) and β-actin (1:2,000) for 4°C overnight. Both antibodies were purchased from Santa Cruz Co. (Santa Cruz, CA, USA). After washing with TBS-T, the membrane was incubated with secondary antibody, and the signals were visualized using the enhanced chemiluminescent detection reagent from Pierce (Rockford, IL, USA).
Cell viability detection
The cell viability of SW1990 and Capan-1 cells with or without eIF3a depletion were assessed using a cell counting kit-8 (CCK-8) assay (Beyotime, Nantong, China) based on the manufacturer’s instructions. Briefly, cells transfected with (shRNA group) or without specific shRNA against eIF3a (negative control group, NC group) were seeded into 96-well plates at an initial concentration of 6,000 cells per well. Cells were incubated for a total of 7 days. On each monitored day, 10 μL of CCK-8 solution was added to each well and cells were further incubated at 37°C for 2 hours, the absorbance was measured by an ELISA reader at a wave length of 450 nm.
Colony formation assay
Anchorage-independent growth was also assessed by colony formation ability. A total of 1,000 cells were seeded in 6 cm-well plates and cultured at 37°C. The culture medium was changed at regular intervals after 10–15 days according to the character of each cell line. The adherent cells were washed with PBS twice, fixed with 4% paraformaldehyde, and stained with 0.1% crystal violet (Sigma-Aldrich, NY, USA). The total number of colonies (≥ 50 cells/colony) was counted, and the results were expressed as the average of three independent experiments.
Wound-healing assay
Pancreatic cancer cells SW1990 and Capan-1 were seeded into 6-well plates and infected with either control or specific shRNAs. Forty-eight hours post-transfection, cells were washed with pre-warmed PBS and then 10 uL sterile pipette tips were used to scrape a straight line in the center of each group. After scratch, cells were rinsed again with PBS for three times and incubated with serum-free medium immediately. In 37°C incubator, cells were allowed to grow for additional 24 hours when scratches were observed and photographed. Images were captured using a Nikon microscope at a magnification of 200 ×. Each assay was repeated at least three times in triplicate.
Transwell assay
SW1990 and Capan-1 cells were cultured in 24-well plates and infected with specific eIF3a shRNA. After 48 hours, cells were harvested with serum-free DMEM media and 150 μL of cell suspension (approximate 3 × 104 cells) was placed in the upper chamber (Corning, New York, NY, USA). Meanwhile, the lower chamber was added with 600 μL DMEM medium supplied with 10% FBS. As for the invasion assay, the chamber was pre-coated with Matrigel (Corning, NY, USA) for 6 hours before seeding cells. After incubated for additional 12 hours, cells were fixed with pre-cold methanol for 10 minutes and then stained with crystal violet (0.1%) for 5 minutes. Afterwards, the images were captured under a microscope (Nikon).
Mouse xenografts
Female Balb/c nude mice (6–8 weeks of age, weighing 18–20 g) were obtained from Shanghai Laboratory Animal Center (Shanghai, China). The mice were housed in micro-isolator cages with autoclaved bedding in a specific pathogen-free facility with 12-hour light/dark cycles. The mice were divided into two groups (NC group and shRNA-injected group, n = 5 for each group). According to body weight before the experiment, 2 × 106/site cell was subcutaneously injected into the right flanks of the nude mice. Tumor growth was monitored every 3–4 days and was measured in two dimensions. The tumor volume was calculated using the following formula: tumor volume = width2 (mm2) × length (mm)/2, where width and length were the shortest and longest diameters, respectively. After four weeks, the mice were sacrificed, and the primary tumors were dissected and measured. All animal experiments were performed in compliance with the Ethics Committee guidelines of the first affiliated hospital, Zhejiang University.
Statistical analysis
Statistical analysis was conducted using SPSS software version 18.0 (SPSS Inc., Chicago, IL, USA). The Pearson χ2 test was used to analyze eIF3a expression in pancreatic cancer and normal pancreatic tissue. The results were presented as the mean ± standard deviation (SD) of three independent experiments. A P value of < 0.05 was considered significant.
RESULTS
Aberrant eIF3a expression in human pancreatic cancer tissues and the eIF3a expression correlated with tumor metastasis and tumor staging
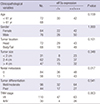
To investigate eIF3a expression level in human pancreatic tissues, qRT-PCR and immunohistochemistry staining assays were performed to detect eIF3a in both normal and pancreatic cancerous tissues. As revealed in Fig. 1A, in the obtained 30 tissues, the relative mRNA level of eIF3a in human pancreatic ductal carcinoma tissues was elevated by seven-fold as compared with that of normal counterpart (P < 0.001). Furthermore, in the slides from 140 cases of pancreatic ductal carcinoma, IHC staining showed that eIF3a was abundantly detected in malignant ductal adenocarcinoma patients, but not in normal pancreatic duct epithelial tissues (Fig. 1B). Overall, aberrant eIF3a expression in malignant lesions of the pancreas was observed. The aberrant eIF3a expression was then statistically analyzed with the clinicopathological variables (Table 1). It was revealed that the expression of eIF3a was not significantly correlated with variables such as age of onset, gender, tumor size, location and differentiation (P = 0.159, 1.00, 0.101, and 0.541, respectively). However, statistical correlation of eIF3a expression with nodal metastasis and TNM stage were observed (P = 0.017 for metastasis, and P = 0.003 for TNM stage). These observations suggested that the overexpresssion of eIF3a was associated with the aggressiveness in pancreatic ductal adenocarcinoma.
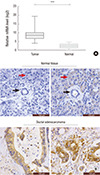 | Fig. 1Aberrant eIF3a expression in pancreatic cancer tissues.
(A) Relative eIF3a mRNA levels in the pancreatic ductal adenocarcinoma tissues (n = 30) and their paired adjacent non-cancerous pancreas tissues (n = 30). (B) Immunohistochemistry analysis of the protein level of eIF3a in slides from normal pancreas tissues and the pancreatic ductal adenocarcinoma tissues (n = 140). It was shown that eIF3a was non-expressed or lowly expressed in normal pancreatic duct (black arrow) and normal pancreatic acini (red arrow). In contrast, eIF3a was strongly expressed in the cancer tissues.
*P < 0.001
|
Table 1
Association between eIF3a expression and the clinicopathological variables in 140 cases of pancreatic ductal adenocarcinoma
![]()
Constitutive eIF3a expression in human pancreatic cancer cell lines and knockdown of its expression in vitro
The high eIF3a expression level in malignant pancreatic lesions prompted eIF3a expression assessment in pancreatic cancer cell lines. In all the seven cell lines that were examined, Western blot analysis detected eIF3a protein levels (Fig. 2A). SW1990 and Capan-1 cells displayed the highest eIF3a protein level, whereas Miapaca-2 cells had the lowest eIF3a expression level. Therefore, SW1990 and Capan-1 cell lines were chosen for the subsequent analyses. Specific shRNA against eIF3a was stably transfected into SW1990 and Capan-1 cells to explore the detailed role of eIF3a in tumorigenicity. The efficiency of eIF3a knockdown was confirmed through Western blot analysis where the protein level of eIF3a was barely detected after transfection of the specific shRNA into SW1990 and Capan-1 cells (Fig. 2B). These data suggested the high specificity and transfection efficacy of eIF3a shRNA.
 | Fig. 2The constitutive expression of eIF3a in pancreatic cancer cell lines and the knockdown efficacy of a specific shRNA against eIF3a. (A) In the seven pancreatic cancer cell lines, it was observed that SW1990 and Capan-1 cells exhibited the strongest expression of eIF3a, whereas Miapaca-2 cell line exhibited the least expression of eIF3a. Hence, SW1990 and Capan-1 were chosen for subsequent analyses. (B) A specific shRNA against eIF3a was utilized to knock down the expression of eIF3a in SW1990 cells and Capan-1 cells. Western blot analysis revealed that the protein level of eIF3a was barely detected after transfection of the specific shRNA into these two cell lines, suggesting the high efficacy of our designed shRNA.
|
Knockdown of eIF3a inhibited cell proliferation and colony formation in pancreatic cancer cells
Next, CCK-8 assay and colony formation assay were performed to explore the detailed role of eIF3a in pancreatic cancer cells. In cell proliferation assay, significant disparities were observed from the fourth day in SW1990 cells and the fifth day in Capan-1 cells (Fig. 3A). The inhibitory effects grew as the time extended in both cells. As for the colony formation assay, it was shown that knockdown of eIF3a caused visual decreases of colonies in both cell lines (Fig. 3B). By counting the colonies numbers, it was further shown that more than 60% of SW1990 colonies and 75% of Capan-1 colonies were suppressed upon eIF3a shRNA transfection (Fig. 3C). These results indicated that knockdown of eIF3a retarded cell proliferation and colony formation in pancreatic cancer cells.
 | Fig. 3Knockdown of eIF3a inhibited cell proliferation and colony formation in pancreatic cancer cell lines. (A) In the cell proliferation assay, significant disparities were observed from the fourth day in SW1990 cells and the fifth day in Capan-1 cells. The inhibitory effects grew as the time extended in both cells. (B) In the colony formation assay, it was shown that knockdown of eIF3a caused visual decreases of colonies in both cell lines. (C) By counting the colonies numbers, it was further shown that more than 60% of SW1990 colonies and 75% of Capan-1 colonies were suppressed upon eIF3a shRNA transfection. Data were obtained in triplicate with each experiments repeated three times.
*P < 0.05; †P < 0.01
|
Knockdown of eIF3a inhibited wound recovery abilities in pancreatic cancer cells
Wound-healing process reflects the cell migration abilities after initial scratches. To assess the wound recovery abilities, both SW1990 cells and Capan-1 cells were subject to wound-healing assays. Twenty-four hours after scratch, it was observed that the wound was mostly closed in the control SW1990 and Capan-1 cells. However, eIF3a-depleted cells barely recovered the initial scratches (Fig. 4, upper panels). Consistently, as compared with the control cells, eIF3a-depleted SW1990 and Capan-1 cells exhibited only approximately 50% migration abilities as reflected by the recovered wound areas (Fig. 4, lower panels). These observations suggested that eIF3a was associated with cell migration abilities in both SW1990 and Capan-1 cells.
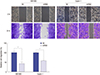 | Fig. 4Knockdown of eIF3a inhibited the wound recovery in SW1990 cells and Capan-1 cells. Both SW1990 cells and Capan-1 cells were subject to wound-healing assays after transfection with scramble or specific shRNA against eIF3a. Twenty-four hours after the scratch, wound recovery rates were photographed and the recovered areas which represented cell migration were quantified and averaged from three independent assays.
*P < 0.01
|
Knockdown of eIF3a decreased cell migration and invasion abilities in human pancreatic cancer cell lines
Moreover, knockdown of eIF3a with specific shRNA inhibited cell migration and invasion based on observations from transwell assays (Fig. 5). The migration rate was inhibited by 55% for SW1990 cells and 46% for Capan-1 cells. Meanwhile, cell invasion rate was decreased by more than 60% for both cell lines. All of these data jointly suggested that eIF3a could promote cell metastasis in human pancreatic cancer.
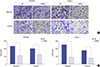 | Fig. 5Knockdown of eIF3a decreased cell migration and invasion abilities in human pancreatic cancer cell lines. (A) SW1990 cells and Capan-1 cells were subject to transwell assays after cells were depleted of eIF3a by shRNA. Visually, the transmigrated cells were significantly decreased in the shRNA group relative to control group. (B) After counting the transmigrated cells, it was shown that the migration rate was inhibited by 55% for SW1990 cells and 46% for Capan-1 cells. Meanwhile, cell invasion rate was decreased by more than 60% for both cell lines. Each assay was repeated for three times.
*P < 0.01
|
Knockdown of eIF3a inhibited the tumorigenic ability in a xenotransplanted model
Two pancreatic cancer cell groups (NC and shRNA against eIF3a) were subcutaneously implanted into the nude mice to determine the impact of eIF3a depletion on pancreatic tumor growth in vivo. As shown in Fig. 6A and 6B, the tumor volume of eIF3a-depleted xenografts was significantly decreased compared with that of tumors formed by scramble shRNA-transfected cells. After four weeks, mice were sacrificed and tumors were all dissected and weighed. The tumors were generally lighter when eIF3a was knocked down in experimental mice (Fig. 6C). These data indicated that eIF3a displayed a potential tumorigenic ability in vivo.
 | Fig. 6Knockdown of eIF3a inhibited the tumorigenic ability in a xenotransplanted model. (A) Capan-1 cells with or without eIF3a depletion were subcutaneously injected into the nude mice (n = 5 for each group) to determine the impact of eIF3a depletion on pancreatic tumor growth in vivo. Tumor volumes from two groups of mice were monitored for a consecutive 4 weeks. (B) After four weeks, tumors were dissected. (C) Tumor weights from each group were weighed. It was shown that the average tumor weight from eIF3a-depleted group was significantly decreased as compared with that in the control group.
*P < 0.05
|
DISCUSSION
Pancreatic cancer still remains a serious health problem, with a 5-year survival rate for all stages at < 5% (1). Most patients were diagnosed at an advanced stage with poor prognosis. Novel molecules crucially involved in pancreatic cancer cell growth are urgently needed for early detection and intervention.
As the largest and most complex subunit of eIF3, eIF3a has been identified in a wide range of eukaryotic organisms, including fungi (21), yeast (22), insects (23), plants (24) and mammals (25). Due to its important regulatory effects in protein translation initiation, it is involved in various cancers, including lung cancer (26) and tumors of stomach (27), urinary bladder(28), colon (29) and ovarian (30). However, the detailed role of eIF3a in human pancreatic cancer remains to be uncovered.
The present study showed that eIF3a was abundantly detected in pancreatic cancer cell lines and malignant ductal adenocarcinoma tissues, but not in normal pancreatic duct epithelial tissues, suggesting the exclusively high expression of eIF3a in cancerous tissues. Initially, the overexpression of eIF3a was observed in breast cancer (31). Later on, the overexpression of eIF3a was also widely reported in other human tumors. Our data together with previous reports collectively suggest the prevalence of eIF3a overexpression in the development of cancers.
Moreover, knockdown of eIF3a in pancreatic cancer cells inhibited cell proliferation and metastasis. While control cells proliferated normally in the culture medium, eIF3a-depleted SW1990 and Capan-1 cells were significantly inhibited from cell proliferation and clonogenisis. Consistently, the pancreatic cancer cell growth rates were also impaired in xenotransplanted mice. Moreover, wound-healing assay showed that depletion of eIF3a significantly slowed down the wound recovery processes in SW1990 and Capan-1 cells. Transwell migration and invasion assays further showed that cell migration and invasion abilities were significantly inhibited by knockdown of eIF3a in SW1990 and Capan-1 cells. All these data strongly suggest that eIF3a is critical for cell proliferation and metastasis in pancreatic cancer.
In addition, we observed the association between eIF3a expression and common clinicopathological variables. The expression of eIF3a in clinical pancreatic cancer tissues were significantly correlated with tumor TNM stage (P = 0.003) and nodal metastasis (P = 0.017). Previously, eIF3a was observed to be associated with patients’ survival in ovarian cancer (19). In non-small cell lung cancer, altered eIF3a predicted the prognosis of this malignancy (26). These clinical studies indicated that the aberrant expression of eIF3a in human cancers might serve as a prognostic factor that could be detected in clinic.
Of particular interest to us, despite wide reports on eIF3a, the pathways that contribute to eIF3a-mediated biological activities remain largely unrevealed. Interestingly, as a translation initiator, eIF3a could affect cancer phenotype independent of translation initiation (20), indicating that translation initiation is not the only pathway that underlies eIF3a function in human cancers. On the contrary, eIF3a is more possible to suppress cellular protection against DNA damages via inhibiting DNA repair, leading to higher frequency of gene mutation for tumorigenesis and lower sensitivity of cancer cells to DNA-damaging-associated anticancer drugs (1516). Hence, eIF3a may function to control DNA repair machinery in the development of human cancers which may be independent of translation initiation. However, more work needs to be done.
In all, the present study identified eIF3a as a critical mediator of cell proliferation, migration and invasion in pancreatic cancer. Knockdown of eIF3a by specific shRNA significantly inhibited cell proliferation and clonogenic abilities in vitro and tumor growth in vivo. High expression of eIF3a correlated with tumor aggressiveness and was closely associated with cancerous cell migration and invasion abilities in pancreatic cancer. Therefore, eIF3a may be a novel target molecule in drug development for pancreatic cancer treatment and prevention. Molecular therapies against eIF3a might be a novel strategy as for the early diagnosis and treatment of pancreatic cancer.
 XML Download
XML Download