

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
X-linked ichthyosis (XLI, OMIM#308100) is a relatively rare hereditary keratinization disorder that affects between 1 in 2000 and 1 in 6000 males worldwide (1). Patients with XLI typically manifest generalized dryness and scaling of the skin with polygonal, thick dark scales that affect the flexures (2), whereas the palms and soles are usually spared. XLI is caused by a partial or complete deletion in the steroid sulfatase gene (STS), which leads to a deficiency of steroid sulfatase (SSase) activity (2). SSase is a 62 kDa microsomal enzyme that is essential for the hydrolysis of 3β-sulphate esters from cholesterol sulfate (CSO4) (3) and a range of other sulfated steroid hormones. Lack of this enzymatic activity results in an accumulation of cholesterol sulfate in the stratum corneum (SC) and an abnormal desquamation of corneocytes, resulting in the typical scaling phenotype exhibited in the disease. The STS gene is located on the distal tip of the short arm of the X chromosome, locus Xp22.3. To detect the deletion of STS gene, multiplex DNA amplification (4), Southern blot (5), and PCR (6) methods have been employed, but more recently, the fluorescence in situ hybridization (FISH) technique has been utilized for a rapid identification of deletions (7). FISH analysis is particularly useful in the diagnosis of carriers, as it can detect genes even if their expression is suppressed (89), though false negatives can occur with point mutations. Recently, the array comparative genomic hybridization (CGH) analysis has been developed to allow high-resolution, genome-wide screening of segmental genomic copy number variations (10). As a result, the diagnosis of XLI has become even more accurate. In addition, other mutations that could modify the XLI phenotype (such as in the filaggrin gene) may also be discovered by array CGH analysis, although the limiting factor is that point mutations cannot be detected.
As the amount of cholesterol declines and cholesterol sulfate accumulates in the intercorneocyte lipids, XLI patients present a mild epidermal permeability barrier abnormality characterized by a delay in recovery (111213). However, other reports describe only a minor basal barrier abnormality in XLI (1415).
Whereas clinical and genetic features of XLI in Caucasian patients are well established, studies in Koreans are lacking, and only a few cases have been reported (1617). In this study, we recruited 16 male Korean patients diagnosed with XLI by FISH and array CGH analysis. We found normal skin permeability barrier function and an unexpectedly frequent kallikrein 7 (KLK7) mutation in Korean XLI patients, which may differentiate them from Western XLI patients.
MATERIALS AND METHODS
Selection of XLI patients and healthy controls, measurement of skin barrier function and sampling
The 16 patients who were enrolled in this study were all diagnosed with XLI by FISH and array CGH analysis. Basic patient information is listed in Table 1. Controls included healthy male individuals between the ages of 19 and 35 with no personal or family history of ichthyosis or atopic diseases.
Table 1
Summary of the clinical features of XLI patients

![]()
To evaluate clinical manifestations of XLI, a questionnaire was given to all prospective patients about their past medical, atopic, and family history, as well as the progress of their skin lesions. For the analysis of skin barrier properties, trans-epidermal water loss (TEWL), SC hydration, and skin surface pH were measured from both lesional and non-lesional skin using Tewameter® (C & K, Cologne, Germany), Corneometer® (C & K) and VARIO pH Meter® (WTW, Weilheim, Germany) respectively. SC samples for the cytokine analysis were collected in the volar forearms by D-squame (CuDerm Corporation, Dallas, TX, USA) stripping. A total of ten consecutive tape strips was taken from the same site. Skin specimens were obtained by punch biopsy taken from the lesional skin of XLI patients and the normal skin of five healthy controls. Hematoxylin-eosin (H & E) and immunohistochemical staining for filaggrin, involucrin, KLK7, and protease-activated receptor-2 (PAR-2) with mouse monoclonal anti-human antibodies (Abcam, Cambridge, UK) were performed on the samples. Blood samples were also taken from the patients for genetic analysis. Two patients (patient 15 and 16) were not able to participate in blood tests and D-squame stripping.
Semi-quantitative analysis of immunohistochemical stains for filaggrin, involucrin, KLK7 and PAR-2
All specimens were analyzed by a dermatologist who was blind to the specimen information. The staining intensity was graded from 0 (negative) to 4 (strongly positive) under a light microscope at a × 200 magnification.
Array CGH analysis
DNA was extracted from peripheral blood using the PureGene kit according to the manufacturer’s instructions (Gentra Systems, Minneapolis, MN, USA). The array CGH chip data were analyzed using the chromofluor image analysis system (Array Analysis; Macrogen, Seoul, Korea). The slides contained 1,440 human BAC clones, including specific loci of more than 40 chromosomal disorders and 356 cell growth related genes from BAC libraries at an average resolution of 2.3 Mb per the entire genome. Each BAC clone was represented on the array in triplicate spots, and each array was scanned using a GenePix4000B scanner (Axon Instruments, Foster City, CA, USA) and analyzed with the array software (MAC VIEWER, Macrogen). Green (test) to red (reference) (G/R) ratios were automatically determined for each sample, and the normalized G/R ratio represented the relative average number of copies of the sequence for those spots that were selected as controls. Spots with G/R ratios higher than the mean plus 2.5 standard deviations (1.25) were considered amplifications or gains of the indicated copy number; and those lower than the mean minus 2.5 deviations (−0.75) were considered losses of the copy number (1819).
FISH analysis with an STS probe
The chromosome analysis was performed according to standard methods using cultured cells from peripheral blood samples obtained from the patients. FISH analysis was performed to confirm array CGH result. In our opinion, these two methods are the best complementation of each other, and thus, no other methods were used. The FISH studies on metaphase spreads of cultured peripheral leukocytes stimulated with phytohemagglutinin were performed using an STS probe (Vysis LSI STS, Abbott Laboratories, Chicago, IL, USA) on Xp22.3.
Cytokine assays
Cytokine assays were performed using the Milliplex® Map Human cytokine Panel I and Panel II kit (Millipore, Bedford, MA, USA) on a Luminex® 200 multiplex system (Invitrogen, Carlsbad, CA, USA). All reagent dilutions (beads, cytokine standards, cytokine controls, biotinylated detection antibody, etc.) were prepared according to the manufacturer’s instructions, and cytokine assays were performed according to the vendor-provided instructions. The patients' corneocyte lysate was diluted 1:2 (25 μL of sample diluent plus 25 μL corneocyte lysate of patient) and incubated for 10 minutes on a rotator.
The cytokine standards, controls, samples, and microspheres were incubated for 2 hours at room temperature on a rotator by using a 96-well filter bottom microtiter plate (Millipore) to allow subsequent washing. This step was followed by addition of 100 μL of biotinylated monoclonal antibodies to each well of the microtiter plate. Following a second 30 minutes incubation on the rotator and washing, 100 μL of streptavidin (10 μg/mL)-conjugated R-phycoerythrin (Invitrogen) were added to each well. After 30 min incubation and a final wash, the microspheres were resuspended in 200 μL of washing buffer, and the 96-well microplate was placed in a Luminex 100 instrument with an XY platform. The amount of cytokine bound to the microspheres by this antibody sandwich technique is determined by the median fluorescence intensity (MFI) of the reporter molecule, phycoerythrin. The MFI of the unknown sample is then converted into a picograms-per-milliliter value based on the known cytokine concentrations of the standard curve by using a five-parameter regression formula. The sequencing was performed by Bioinfra. Inc. (Bioinfra Inc., Seoul, Korea).
Genetic analysis by PCR-Reverse Blot Hybridization Assay (PCR-REBA)
To prepare the DNA template from SC samples, DNA extraction was performed with the QiaAmp DNA mini kit according to the manufacturer’s instructions (Qiagen, Hilden, Germany).
REBA test (M & D, Wonju, Korea) was based on the reverse blot hybridization principle. PCR was performed using a 50 μL reaction mixture containing 2 × master mix, 1 × primer mixture as specified by the manufacturer’s instructions (Genetbio, Daejeon, Korea), and 5 μL of sample DNA. Sterile distilled water was added to yield a final volume of 50 μL. For the REBA test, the first 10 PCR cycles comprised initial denaturation at 95°C for 30 seconds, followed by annealing and extension at 60°C for 30 seconds. These 10 cycles were followed by 40 cycles of denaturation at 95°C for 30 seconds, followed by annealing and extension at 54°C for 30 seconds. After the final cycle, samples were maintained at 72°C for 7 minutes to complete the synthesis of all strands. The amplified target was visualized as a single band corresponding to a length of 100–200 bp by the Chemi Doc system (VILBER LOURMAT, Mare-la-vallée, France). The 5′ ends of all reverse primers were labeled with biotin for chromogenic detection. The amplified PCR products were subjected to reverse blot hybridization assay. A series of oligonucleotide probes (5’ amino labeled) based on KLK7, serine peptidase inhibitor, Kazal type 5 (SPINK 5), or filaggrin (FLG) genes were synthesized and then selected with PCR products from blood samples. Hybridization and washing were performed according to the manufacturer’s instructions. In brief, the biotinylated PCR products were denatured at 25°C for 5 minutes in a denaturation solution (0.2 N NaOH, 0.2 mM EDTA) and diluted in 970 μL hybridization solution (HS; 2 × SSPE/0.1%SDS) before incubation with the REBA membrane strip in a provided blotting tray. Denatured single-stranded PCR products were used to hybridize with the probes on the strip at 50°C for 30 minutes. Primers and probes used are listed in Tables 2 and 3. The strips were then washed twice with gentle shaking in 1 mL of 2 × SSPE/0.5% SDS for 10 minutes at 62°C, incubated at 42°C with a streptavidin-alkaline phosphatase conjugate diluted 1:2,000 (Roche, Grenzach-Wyhlen, Germany) in 2 × SSPE/0.5% SDS for 30 minutes, and washed twice with 1 mL TBS pH 7.5 (0.05 M Tris, 0.15 M NaCl) at room temperature for 1 minute. The colorimetric hybridization signals were visualized by adding alkaline phosphatase-mediated staining solution diluted 1:50 NBT/BCIP, Nitro blue tetrazolium chloride and 5-Bromo-4-chloro-3-indolylphosphate, toluidine salt in 67% DMSO [(v/v)], (Roche) and incubated until the color was detected. Finally, the band pattern was read and interpreted. After testing by REBA, amplified DNA was confirmed by sequencing with an ABI 3730 automated DNA sequencer, using the ABI Prism BigDye Terminator (Cosmo, Ulsan, Korea). The sequences obtained were compared with sequences in the GenBank database for species assignment.
Table 2
Primers used for PCR amplification

![]()
Table 3
The probes used in the PCR-REBA method

![]()
Statistical analysis
To compare the TEWL, SC hydration, skin surface pH and SC cytokine expression levels between the patient and control groups, we performed the nonparametric Mann-Whitney statistical analysis. The paired t-test was used to determine the significance between lesional and non-lesional skin. Statistical analyses were performed using Prism 5 (Graphpad Software, San Diego, CA, USA). The Mann-Whitney test was used for the comparison of the immunohistochemical staining intensity between XLI patient and healthy control groups. Pearson’s chi-square test was done to analyze the statistical difference in frequency of barrier-related gene mutations between XLI patient and healthy control groups (SPSS for Windows, Version 18.0, SPSS Inc., Chicago, IL, USA).
RESULTS
Array CGH analysis and FISH
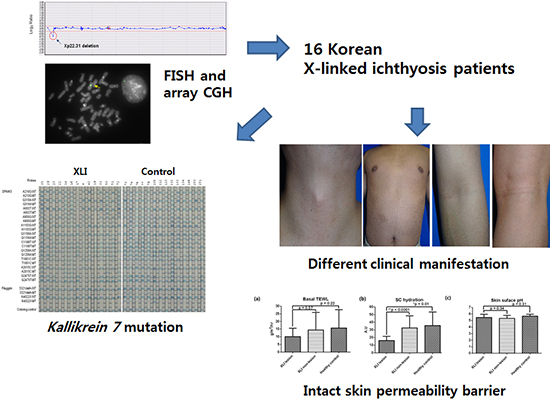
We performed a whole genome array CGH using CGH array slides containing 1,440 clones with 356 cell growth-related genes from BAC libraries. We also simultaneously performed FISH to confirm abnormal array CGH results. Array CGH showed an approximately 0.6 Mb deletion on chromosome Xp22.31 [arr[hg19] Xp22.31p22.31(7,078,532-7,676,445) × 0]. In order to identify where in the segment of chromosome Xp22.31 the deletion was, FISH with a Xp22.31 region specific probe was performed according to the manufacturer’s instructions. We confirmed a deletion in the Xp22.31 region by FISH analysis [46,XY.ish del(X)(p22.31p22.31)(STS-)] (Fig. 1).
 | Fig. 1Deletion in chromosome X. (A) Array CGH results for chromosome X. Arrow indicates the deletion of the steroid sulfatase deficiency critical region (Xp22.31), including the STS gene (arr[hg19] Xp22.31(7,078,532-7,676,445) × 0). (B) FISH with a Xp22.31 region-specific probe; arrow indicates a deletion of the probe (STS-) in a del(X)(p22.31p22.31) chromosome.
|
Skin barrier properties
We compared basal skin barrier function, SC hydration, and skin surface pH in lesional and non-lesional skin from patients, as well as in healthy controls. Whereas basal TEWL levels did not differ significantly among these groups, SC hydration was significantly lower in lesional than in non-lesional or control skin. SC hydration of the non-lesional skin from patients was lower than that measured in the control group, although this result was not statistically significant. Finally, skin surface pH did not differ among the lesional and non-lesional skin of patients and the skin of healthy controls (Fig. 2).
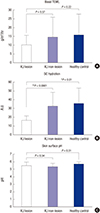 | Fig. 2Skin barrier properties of XLI patients. (A, B) Whereas basal TEWL levels do not differ significantly among these groups, SC hydration is significantly lower in lesional than in non-lesional or control skin. (C) Skin surface pH does not differ among lesional, non-lesional skin of patients and that of normal controls.
|
Mutations in skin barrier-related genes
The results of PCR-REBA performed on patient samples are presented in Fig. 3 and Table 4. Out of 14 patients who provided samples for the analysis, seven (50%) showed a polymorphism in the KLK7 gene (mutant type; MT), other five (35.7%) presented with a mixed type (WT-MT mix) and two (14.3%) were wild type (WT). None from the age-matched healthy control group showed a polymorphism in KLK7 (MT), and 12 controls (85.8%) showed a mixed type. Significantly more of the XLI patients had a polymorphism in the KLK7 gene (MT) as compared to the age-matched healthy control group (P < 0.05) (Table 5). None of the patient or control groups showed mutations in FLG gene, whether 3321delA or K4022X, which are the most frequent FLG mutation (over 95%) found in Korean atopic dermatitis (AD) patients. There were no significant differences in the frequency of mutations in the SPINK5 gene between the patient and the healthy control groups.

Table 4
Results of the PCR-REBA performed in XLI patients

![]()
Table 5
Results of the genetic analysis

| MT | WT-MT mix | WT | P value | |
|---|---|---|---|---|
| XLI patients | 7 (50%) | 5 (35.8%) | 2 (14.2%) | 0.007 |
| Healthy controls | 0 (0%) | 12 (85.8%) | 2 (14.2%) |
Significantly more XLI patients had a polymorphism in the KLK7 gene compared to age-matched healthy control group (P < 0.05).
MT, mutant type; WT-MT mix, mixed wild type and mutant type; WT, wild type.
![]()
Histopathological findings and semi-quantitative evaluation of filaggrin, involucrin, KLK7 and PAR-2 expression
The histopathological findings from the H&E staining of all patients showed hyperkeratosis and a normal to a slightly increased granular layer in the epidermis, but no acanthosis (Fig. 4A and 4B). Immunohistochemical staining for filaggrin and involucrin in the epidermis showed a slightly increased pattern in XLI patients compared to the healthy controls (Fig. 4B-4F). The staining intensities of KLK7 and PAR-2 were significantly higher in XLI patients than in the control group (Fig. 5A-5E). When comparing XLI patients with and without the KLK7 polymorphism, we found that expression of KLK7 did not show any significant differences, whereas expression of PAR-2 was significantly higher in XLI patients with the KLK7 polymorphism (Fig. 5F-5J).
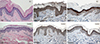 | Fig. 4Histopathological findings. (A-C) XLI patients. (D-F) Healthy control group. H & E staining from all patients shows hyperkeratosis and a normal to a slightly increased granular layer in the epidermis, but no acanthosis. Immunohistochemical staining intensity of filaggrin and involucrin shows a slightly increased pattern in the patient as compared to the control group (magnification 200 ×).
|
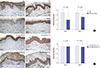 | Fig. 5Immunohistochemical stains and semi-quantitative expression analysis. (A, B) KLK7 and PAR-2 in XLI patients. (C, D) Control group. The staining intensity score is (A) 3, (B) 3, (C) 0.5, and (D) 2. (E) Staining intensities of KLK7 and PAR-2 are significantly higher in XLI patients than in the control group. (F, G) KLK7 and PAR-2 in XLI patients with KLK7 polymorphism, (H, I) without the KLK7 polymorphism. The staining intensity scores are (F) 4, (G) 4, (H) 3, and (I) 3. (J) Expression of KLK7 is not significantly different, whereas expression of PAR-2 is significantly higher in XLI patients with the KLK7 polymorphism (KLK7, 200 ×; PAR-2, 200 ×).
|
Cytokine expression in the SC
We analyzed cytokine expression levels in the SC obtained from the lesional and non-lesional skin of XLI patients, and compared them with values in healthy controls to investigate if inflammation is present in the skin. The cytokines we analyzed included tumor-necrosis factor (TNF)-α and IL-1α, β, which are known as pro-inflammatory cytokines. There were no differences in the expression level of these cytokines among the lesional and non-lesional skin of XLI patients and the healthy control skin (Fig. 6).

Clinical manifestations
The clinical features of XLI patients included in this study are summarized in Table 1. Among the 16 patients, six (37.5%) had their skin lesions present at birth, three (18.8%) by 1 year of age, but seven (43.7%) reported that skin lesions only appeared after their first year of life. Over time, five patients (31.3%) described an improvement in skin symptoms, seven (43.8%) observed no change, and four (25.0%) noted that their skin lesions became worse with age. Thirteen patients (81.3%) lacked involvement of the flexural skin, whereas only three patients (18.8%) exhibited involvement of the flexures. Fig. 7 shows the thick polygonal scales on the trunk, and the lack of flexure involvement in one of the cases.
 | Fig. 7Skin findings from an XLI patient (Case 6). (A, B) The patients in our cohort manifested thick polygonal scales on the trunk. (C, D) Most (81.3%) of the patients lacked flexure involvement.
|
Nine patients (56.3%) described a past history of atopic diseases, including AD or allergic rhinitis and asthma, whereas seven patients (43.8%) reported no atopic history. In the part of the questionnaire relating to the family history of atopic diseases, only four patients (25.0%) reported a positive family history, whereas 12 (75.0%) had no family history of atopic diseases.
DISCUSSION
In the epidermis, SSase hydrolyzes 3β-sulfate esters from CSO4. In XLI, the SSase activity is absent, and CSO4 accumulates and concentrates in the SC interstices. It is reported that the higher proportion of CSO4 in the epidermis alters the physical properties of SC and increases SC intercellular cohesion (20). Previous studies on the skin permeability function of XLI patients showed variable results, but it has been generally accepted that SC integrity and permeability barrier function are mildly impaired (212223). This implies that increased CSO4 may play a role in the disruption of the permeability barrier observed in XLI patients. A recent study by Groen et al. (24) reported that a model for the lipid composition similar to XLI, achieved by addition of CSO4, exhibited increased permeability. However, the barrier abnormality was thought to also be caused by decreased cholesterol in the SC. Decreased cholesterol alone can produce abnormal extracellular lamellar membranes. Therefore, the abnormal skin barrier function in XLI was reportedly due to lamellar/non-lamellar phase separation resulting from excess CSO4 and reduced cholesterol content of SC lamellar membranes (25). Additionally, XLI manifests with delayed desquamation and delayed degradation of corneodesmosomes (CDs), which results in retention hyperkeratosis (25). Interestingly, CSO4 was known to be a serine protease inhibitor (25), and KLK7 (stratum corneum chymotryptic enzyme; SCCE) as well as KLK5 (stratum corneum tryptic enzyme; SCTE) are two key serine proteases that degrade CDs in vitro (26). These enzymes exhibit neutral pH optima, but the pH of the SC was previously reported to be lower in XLI patients than in unaffected individuals (27). Consequently, serine protease activity is inhibited in XLI skin resulting in the appearance of hyperkeratosis of the retention type.
However, Korean XLI patients examined in our study did not demonstrate an impaired skin barrier function. This was evidenced by no differences being observed in basal TEWL and skin surface pH between the patients and the healthy control group. As we expected, SC hydration of the lesional skin was significantly lower than that of the non-lesional skin or the skin from the healthy control group. SC hydration of the non-lesional skin of patients showed no significant differences when compared to that of the control group. The histological findings of the samples taken from patients showed retention hyperkeratosis, but no acanthosis or increased stratum granulosum, which may appear as a homeostatic response to a defective skin barrier. In addition, the expression levels of pro-inflammatory cytokines, including IL-1α, 1β, and TNFα that are known to be increased in SC when the barrier is disrupted, did not differ in patients as compared to the control group. This result is concomitant with the study by Hoppe et al. (23) showing that no inflammatory or repair mechanisms are triggered in XLI skin, although the patients in their study demonstrated increased basal TEWL. Moreover, in accordance with a previously published study from our group, XLI patients showed normal levels of pyrrolidone carboxylic acid and caspase-14 expression in the SC (28).
As previous reports are based mainly on Western populations, the different features of XLI patients may be attributed to ethnic differences. Since some reports may have included patients diagnosed by clinical assessment and not by detection of the STS genetic mutation, it is possible that some these patients may have had another subtype of ichthyosis, such as ichthyosis vulgaris (IV), but were misdiagnosed owing to clinical similarities. Our study specifically recruited only the patients diagnosed with XLI by FISH and array CGH analysis
Thus far, almost 40 FLG mutations have been reported worldwide (29). However, it is also widely accepted that there are remarkable ethnic differences in the prevalence and specific types of FLG mutations. In the Korean population, the 3321delA and K4022X FLG mutations were reported (3031) in more than 95% of reported mutations despite the studies being conducted in a large AD group with a much lower FLG mutation frequency (6.45%) as compared with European populations. Therefore, we only assessed these two FLG mutations in our patient group. Not only Koreans, but also Chinese (30%) and Japanese (20%) were also reported to have a lower frequency of FLG mutations (303233). As the prevalence of a FLG mutation is much higher in European populations, there is a chance that FLG mutations may have accompanied some of the XLI patients and further impaired their skin barrier function. Additionally, XLI seems to be more common in the Korean population than previously expected. An accurate investigation of the prevalence of XLI and IV in Asian populations is highly desirable. This and other recent FLG mutation studies, indicate that, at least in Korea, many dermatologists and pediatricians have misdiagnosed XLI as IV because they derive their diagnostic criteria from textbooks written in English and based on European or American populations.
The KLK7 gene polymorphism in our patient group was significantly higher than in the healthy control group, whereas there are no previously published studies regarding the KLK7 gene polymorphism in XLI. The polymorphism of the KLK7 gene has been reported in a British AD cohort (34), and it is thought to result in a gain of function of the protease (3435) that causes premature degradation of CDs and consequently a defective skin barrier function. According to Hansson et al., transgenic mice with human SCCE overexpression developed features of inflammatory skin diseases such as AD (36).
Based on our genetic study, we suggest a compensation theory for the KLK7 gene polymorphism observed in our XLI patients. As retention hyperkeratosis due to failure of degradation of CDs by proteases occurs in XLI, a countervailing effort to upregulate the expression of serine proteases may have arisen evolutionarily. Owing to increased expression of KLK7, KLK5 will also be activated by positive feedback loops involving KLK7, and SCTE (KLK5) would in turn activate PAR-2 and increase its production (37).
We were able to demonstrate the increased expression of KLK7 and PAR-2 in XLI patients compared to the control group by immunohistochemical staining. However, KLK7 staining intensity did not differ between patients with and without the KLK7 polymorphism. Additionally, no significant differences in the SC thickness could be seen, although it did appear slightly thicker in the group without than in the group with the KLK7 polymorphism (data not shown). KLK7 is activated only in neutral pH and PAR-2 is known to be activated and exert its role when the pH of SC increases after skin barrier perturbation (3839). Evolutionarily, the expression and production of KLK7 and PAR-2 might be increased to compensate for the retention hyperkeratosis but their actual activity may be inhibited by increased cholesterol sulfate, which also acts as a serine protease inhibitor (40), or by the normally acidic SC pH found in XLI patients. It is possible that other mechanisms increasing KLK7 activity, even in the absence of the KLK7 polymorphism, exist to compensate for retention hyperkeratosis. There were no significant differences in clinical features, basal TEWL, SC hydration and skin surface pH between XLI patients with and without the KLK7 polymorphism. Therefore, a correlation between the KLK7 gene polymorphism and clinical manifestation is yet to be established through further studies.
In this study, clinical features, as well as properties of and genetic mutations related to the skin barrier function were investigated in XLI patients diagnosed with an appropriately sensitive method. Interestingly, we found several novel features that differed from previous studies. In contrast to those reports, XLI patients in our group had skin lesions before, at the beginning, and after the first year of life in 62.5% of cases. The lesions tended to improve with age in 31.3% of patients, and the flexural skin was spared in 81.3% of those affected. In addition, 62.5% of the patients had atopic diseases in the past or in their family history.
To our regret, identification of the actual size of the deletion in an individual patient was impossible with the use of our array CGH analysis, as a BAC chip clone instead of an oligoarray was used in our method. However, the approximate deletion size of all the patients confirmed as XLI was 5,979,143 kb (0.6 Mb), and the location of deletion was between 7,078,532 and 767,644, which is within the STS gene locus. Therefore, we could ascertain that the patients had deletions in the critical region that contains the STS gene. Another limitation of our study was that zymography to evaluate the enzymatic activity of KLK7 could not be performed owing to limited sample material. Instead, we tried to determine the staining intensity by immunohistochemistry. In future studies, a direct measurement of enzymatic activity of KLK7 in a larger cohort will be necessary. However, the sample size of our study is relatively small, as the prevalence of XLI is very low. We have recruited as many subjects as possible for a period of over four months and increasing the sample size further was impossible.
In summary, Korean XLI patients showed different features as compared with previously known characteristics of Western XLI patients. Their skin barrier function is not impaired and they have a significant KLK7 polymorphism involvement, which advances a new compensation theory.
 XML Download
XML Download