

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Filaggrin, a histidine-rich cationic protein that is a decomposition product of profilaggrin, is one of the major components of keratohyaline granule. The name filaggrin (filament-aggregating-protein) is derived from its nature to aggregate keratin filaments (1). Within the stratum corneum, filaggrin monomers become incorporated into the lipid envelope, responsible for the skin barrier function. Alternatively, these proteins interact with keratin intermediate filaments. Filaggrin undergoes further processing in the upper stratum corneum to release free amino acids as natural moisturizing factors (2). Trans-urocanic acid, formed from histidine released from filaggrin, absorbs ultraviolet radiation and protects against thymine dimer formation in keratinocytes. Pyrrolidone-5-carboxylic acid (PCA) is the other main breakdown product of filaggrin and together these organic acids help to maintain the pH gradient of the epidermis as evidenced by a higher surface pH in filaggrin null mutation carriers. The 'acid mantle' of the stratum corneum has a well-known antimicrobial effect (3).
The profilaggrin/filaggrin gene (FLG) resides on chromosome 1q21 and consists of three exons. Exon 3 is extremely large (> 12 kb) and encodes most of the profilaggrin polypeptide with 10-12 repeats, which are almost completely homologous (4). There is a unique “repeat number” polymorphism of FLG, related to the above-mentioned repeats. FLG encodes 10 highly homologous and only slightly genetically different FLG units, but the exact number encoded can vary, with individuals harboring 10, 11, or 12 monomers. The differences are attributable to possible repeats of subunits 8 or 10 or both. It has been suggested that the number of FLG repeats can relate to a dry skin phenotype; i.e., fewer repeats may lead to less FLG protein expression and drier skin (56).
Mutations in FLG have recently been identified as the cause of the common genetic skin disorder ichthyosis vulgaris (IV; OMIM no. 146700), the most prevalent inherited disorder of keratinization. The main characteristics of IV are xerosis, scaling, keratosis pilaris, palmar and plantar hyperlinearity, and a strong association with atopic disorders (78). Interestingly, a strong association between these FLG null alleles and atopic dermatitis (AD) has also been firmly established (9).
Additional studies demonstrated that identification of FLG null alleles is an indicator of a poor prognosis in AD, predisposing patients to a form of eczema that starts in early infancy and persists into adulthood (1011). Highly significant association of the FLG null mutations with eczema and concomitant asthma have been replicated. Moreover, several studies reported that these mutations predispose carriers to asthma, allergic rhinitis, and allergic sensitization in the presence of eczema (12). Recent studies hypothesized that the damage to the skin barrier caused by FLG mutations allows allergens to penetrate into the epidermis and subepithelial tissues and to interact with antigen presenting cells, known as Langerhans cells and dermal dendritic cells, which might further initiate the Th2 immune response and lead to the development of systemic allergies, including allergic rhinitis and atopic asthma (13).
Interestingly, there is significant ethnic differences in the FLG mutation. Akiyama reported most FLG mutations are specific to each population, such as European, Japanese, Singaporean Chinese and Taiwanese (14). Major differences exist in the spectra of FLG mutations observed between different ancestral groups. Prevalent FLG mutations are distinct in both the European and the Asian populations. Only two mutations, R501X and E2422X were found in both European and Asian populations among 27 reported FLG mutations (14).
There are more differences between Japanese and Chinese than there are similarities among Asians (15). So, it is likely there will be different FLG polymorphisms in Koreans. However, currently, there are only a few studies about FLG polymorphisms in Korea, and systematic studies have not been carried out (16). Therefore, we have identified FLG polymorphisms in Korean AD patients using overlapping long-range PCR.
MATERIALS AND METHODS
Study population
The study was made up of three groups including 9 IV patients, 50 AD patients and 55 normal controls. IV was diagnosed by biopsy and physical examination, and AD was diagnosed on the basis of a skin examination by experienced dermatologists, using the criteria of Hanifin and Rajka.
Blood sampling and extraction of genomic DNA
We took participants' blood samples in EDTA tubes and processed them immediately or after refrigeration at a temperature of 4°C. We used G-DEX™ IIb genomic DNA extraction kit (iNtRON Biotechnology, Seongnam, Korea) for extracting genomic DNA from whole blood, and proceeded as follows.
We centrifuged at 2,000 g for 5 minutes after mixing 300 uL whole blood with 900 uL RBC lysis buffer, and then discarded the supernatant, leaving just a small amount of the cell pellet. We shook well causing the karyocytes to float. After adding 300 uL cell lysis buffer and 1.5 uL RNase A solution, we soaked the tubes in a 37°C water bath. Once the tubes returned to room temperature, we added 100 uL protein precipitation buffer and centrifuged at 2,000 g for 5 minutes. Using the supernatant, we made a genomic DNA pellet by inverting the container gently with 300 uL 100% isopropanol. After centrifuging at 2,000 g for 3 minutes, we discarded the supernatant, leaving the genomic DNA pellet. We then resuspended the pellet by gently inverting the container with 300 uL 70% ethanol and centrifuged again. After discarding the supernatant and drying the pellet for 10-15 minutes, we dissolved the dried genomic DNA pellet with 50 uL DNA rehydration buffer and stored at -20°C.
The purity of the DNA was checked by 260/280 optical density using a Nanodrop ND-1000® Spectrophotometer (Thermo Fisher Scientific, Wilmington, MA, USA).
Polymerase chain reaction (PCR)
We basically followed the recently reported “overlapping long-range PCR” method (17), but we transformed some of the experimental conditions. TaKaRa LA Taq™ polymerase (Takara Shuzo Co. Ltd., Shiga, Japan) was used as the DNA polymerase for PCR reactions. PCR reactions using GeneAmp PCR system 2700 (Applied Biosytems, Princeton, NJ, USA) were processed under the following conditions: 1U LA Taq™ polymerase, 500 ng genomic DNA, 10 pmol forward/reverse primers, 0.5 uL 2.5 mM dNTP, 2U 10X LA Taq buffer II (Takara Shuzo Co. Ltd., Shiga, Japan), and 2 uL 5X BD (Solgent Co. Ltd., Daejeon, Korea) were mixed, and 3X distilled water was added to a final volume of 20 uL. Amplified PCR product was observed with electrophoresis in 0.8% agarose gel.
We modified each PCR reaction because each amplified genomic DNA fragmentation used different PCR primers. Each PCR reaction is shown in Table 1.
Table 1
PCR primers used for FLG mutation analysis

DNA sequencing
After amplified genomic DNA fragments were purified with the QIAquick PCR purification kit (Qiagen, Valencia, CA, USA), sequences were determined by an Applied BioSystems 3100/3700 DNA sequencer (Applied Biosytems). Reaction conditions for DNA sequencing were as follows: after denaturing at 96°C for 1 minute, and 25 cycles of 96°C for 10 seconds, 50°C for 5 seconds and 60°C for 4 minutes, the reaction was finalized at 10°C for 10 minutes. Several forward/reverse sequencing primers were used, depending on the size of Genomic DNA fragment.
Statistical analysis
We conducted χ2 test for confirming the frequency differences of genotypes and alleles between AD patients and normal controls, and to determine clinical relevance. Student's t-test was conducted for checking the differences in biophysical measurements. We judged that P < 0.05 was statistically significant in all cases.
RESULTS
Establishing the methodology of FLG DNA sequencing
We tried to follow the reported “overlapping long-range PCR” method for confirming FLG polymorphisms and mutations, but modifications were necessary in most reaction conditions. Annealing temperature was changed for optimum conditions in numerous PCR reactions, and frequently, a 2nd PCR reaction was required to amplify a second band. We summarized these in Table 1.
We established the reaction conditions of PCR and sequencing for most DNA fragments through trial and error; particularly problematic were exon 3 (1-3) and exon 3 (7-10). Exon 3 (1-3) was the longest DNA fragment with 3,697 bp. We got a single-band product with suitable size by PCR reaction, but sequencing did not progress smoothly. For this reason, we did sequencing analysis by dividing exon 3 (1-3) into two fragments, exon 3 (0-1; 1,710 bp) and exon 3 (0-3; 2,462 bp). In the case of exon 3 (7-10), the PCR reaction with the reported forward & reverse primers did not work well. With rescaled primers, we produced the desired PCR product and used it to conduct sequencing analysis (Fig. 1).
Fig. 1
Strategy used in this study for mutation detection for FLG. Newly designed DNA fragments are indicated by red color. As shown in the results of electrophoresis, most DNA fragments were successfully amplified.
With shotgun method, Sasaki et al. reported the presence of distinct copy-number variants in Japanese (designated as A, B, and C), and distinct differences in single nucleotide polymorphism (SNP) among them (18). We could detect A, B, C variant which had been reported in Japan, but could not detect Bs variant with repeat number 10. Meanwhile, we identified new subtype, which had similar SNP pattern with A variant but had 11 repeat numbers with one more repeat on repeat 8. We called it A'. A' have 11 repeat number like C variant but had similar SNP pattern with A variant, so we could divide A' and C by comparing sequence.
FLG polymorphism in IV, AD patients
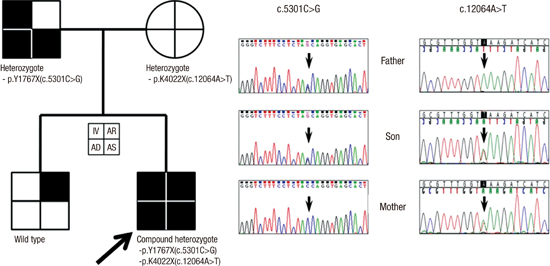
We studied 9 IV patients and their families. Among the 8 families, one family included both father and son with IV, AD and allergic asthma. We checked FLG sequencing in both father and son, and detected a nonsense mutation, p.Y1767X (=c.5301C>G) which confirmed previously in Korean IV patients (Fig. 2) (19). The mother in this family was heterozygous for p.K4022X (=c.12064A>T), which had been reported in Japan (20). The son, as an index case, also had p.K4022X, so he was a compound heterozygote with two simultaneous mutations. The mother of this index case uniquely had no allergic disease, including AD, as well as no IV.
Fig. 2
Novel mutation p.Y1767X in Korean ichthyosis vulgaris family. Also note the known mutation, p.K4022X, in the mother and the index case. Each quadrant was assigned to represent phenotypes.
IV, ichthyosis vulgaris; AD, atopic dermatitis; AS, asthma; AR, allergic rhinitis.

Another index case, a female with AD and allergic rhinitis, was homozygous for p.K4022X. Though her younger brother was also homozygous for p.K4022X, he didn't have IV and AD, but did have severe allergic rhinitis.
We detected no more mutations in other families, but we could confirm 120 SNPs by comparison with the FLG reference sequence (FLG RefSeqGene; NG_016190.1), and 63 of them were unreported new SNPs (Supplementary Table 1). Repeat number polymorphisms were related with 105/120 SNPs. It deemed as expressing genetic character of Korean FLG, because among remnant 15 SNPs, c.10783(T>C) in repeat 10 and 7th (A>G), 310th (C>T), 697th (C>G) in repeat 10.2, etc. included 100% minor allele of FLG reference sequence.
We conducted full sequencing of FLG in 50 AD patients and 55 normal control. About 120 SNPs of IV patients, we compared if the frequency of genotype and allele would be different between AD patients and normal control (Table 2). The results show there were many statistically different SNP loci, and these loci, except c.975G>A, were most concerned with repeat number polymorphism. Bases of variant B and C were especially same and these were seem to be statistically different with variant A and other loci; for example, in case of A/B/C=G/T/T repeat number variant-specific polymorphism such as c.995G>T locus). These pattern was observed through nearly whole 3rd exon (Supplementary Table 1).
Table 2
Comparison of FLG repeat number polymorphism between normal control and atopic dermatitis patients

To compare Japanese data, A' was incorporated into A. Numerics in parentheses denote the frequency of corresponding genotype or allele.
*Adapted from Sasaki et al., J Dermatol Sci 2008 (18).
Loss of function mutations of FLG in AD patients were as follows: there was one heterozygote each of p.Y1767X and p.K4022X, and one compound heterozygote in an IV family with AD and asthma, as shown previously. There were two homozygotes of p.K4022X in a family with AD and rhinitis. We checked mutations for other AD patients and normal controls, and confirmed one more heterozygote of p.K4022X in an AD patient.
In summary, we found two p.Y1767X and five p.K4022X in AD patients and their families (one of them was a compound heterozygote). The frequencies of all participants were 1.6% for p.Y1767X and 4.3% for p.K4022X. Interestingly, p.K4022X was found in 3 patients and 2 normal controls, while p.Y1767X was found only in AD patients. Table 3 shows the clinical characteristics of patients with mutations and their families.
Table 3
Clinical characteristics of carriers of FLG mutation

DISCUSSION
AD is not a simple genetic disease but a multicausative disease (21). Therefore, assuming that IV is a monogenic disease with an effect on AD susceptibility, we have studied 9 Korean families with IV and identified 2 FLG mutations. Our research plan included full sequencing of FLG, considering that there was no systematic study of FLG in Korea and there were obvious differences between ethnic groups in previously reported studies. In addition, we have shown that these genes would be associated with genetic predisposition to AD in Korea.
In comparison to other sequencing methods, the overlapping long-range PCR used in this study was more economical in time and cost, but was limited in its ability to produce consistent PCR products. Therefore, thorough research was needed to confirm the decoded sequence. We processed a minimum of two courses of sequencing from two forward/reverse directions, and a maximum of 6 reads of some overlapping sequences. We confirmed whether decoded DNA fragments were correct by checking well-known SNP loci of FLG. filaggrin repeats are mostly homologous, but the human filaggrin gene (FLG, NG_016190.1) has some variation among repeats, unlike the mouse filaggrin gene (Flg, NC_000069.5) (22). After designing suitable PCR primers, we amplified DNA fragments under optimum conditions and processed them rigorously several times to confirm the sequences. Therefore, we were able to establish a method to sequence the full length of FLG, overcoming previously mentioned limits.
Recent study showed that the percentage of mutations in the FLG gene was 74% and 43% in patients with isolated IV and patients with AD-associated IV, respectively (23). Although we detected no more mutations in other families, but we could confirm 120 SNPs by comparison with the FLG reference sequence. The frequency of mutation is very less while that of SNP (as it is considered polymorphism) is relatively high, and all types of SNPs can have an observable phenotype or can result in disease.
The characteristics of the Korean FLG loss-of-function mutation are as follows: First, the mutation was found primarily in families with personal and familial histories of IV and atopic diseases. It is expected that there will be additional detection of mutations through concentration on research of families with familial histories in the near future. Second, a mutation, p.Y1767X, was related to early onset and severe symptoms, which last until adulthood, and severe AD with other atopic diseases, such as asthma. This suggests that loss-of-function by this mutation caused decreased production of filaggrin and abnormal function of the skin barrier, thus the typical progression of AD, with early onset, long standing and respiratory atopy, might be explained. Third, p.K4022X, showed a different pattern than p.Y1767X. Two of five carriers (mother of p.Y1767X index case, brother of p.K4022X index case) have no history of IV or AD. The mother, as a heterozygotic carrier of p.K4022X, did not have any allergic disease, including allergic rhinitis. Previous research in Japan showed that p.K4022X, although located almost at the end of the FLG 3rd exon, caused problems in post-translational processing and resulted in loss of normal function. They contended that this mutation contributes like the other mutations located in the beginning of the gene (20). However, in our study, we demonstrated that some carriers of p.K4022X had no symptoms of AD and IV and a homozygous index female patient with AD showed a different disease pattern (late onset, mild symptom, etc.). We expect that clinical significance of p.K4022X would be confirmed through an additional and larger study.
Following the article reporting methodological breakthrough on the full-sequencing of the gene encoding FLG, associations between loss-of-function mutations of FLG and atopic dermatitis were reported across ethnicities (14). However, both the low prevalence of FLG mutations in AD patients in some nation (< 4% in Italian) and high prevalence of FLG mutations in healthy control in other nation (~10% in Irish) suggest that factors other than FLG mutation may be at-work (424). So we further conducted analysis of copy number variation (CNV) in Koreans, in order to examine the role of CNV in Korean AD patients.
In conclusion, by using FLG sequencing techniques confirmed in this study, new mutations or genetic polymorphisms with ethnic characteristics would be detected and further larger studies of repeat number polymorphisms could be performed. New biomarkers related to prognosis and treatment can be discovered by the fusion of research of clinical and genetic data. While pharmacologic interventions that directly target filaggrin are a long way from clinical application, personalized medicine may be possible in the future with rapid genetic testing for filaggrin mutations and SNPs.
 XML Download
XML Download