

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Preeclampsia, a disorder with 5%-7% prevalence among pregnant women affects both the mother and fetus and is the main cause of maternal mortality. Although numerous researches have been focused to depict pathogenesis of preeclampsia the exact etiology is not clearly known (1). Currently, a decrease in the uteroplacental perfusion due to inadequate cytotrophoblast invasion of spiral arterioles has been postulated to be the initiating factor in preeclampsia (2, 3). The resulting placental ischemia/hypoxia may stimulate the increase of apoptotic microparticles and the release of placental factors, including inflammatory cytokines and other factors, such as lipid peroxides, reactive oxygen species, soluble fms-like tyrosine kinase and soluble endoglin, into maternal circulation (4, 5).
Recent advances in research technology like proteomics can provide valuable information on dynamic changes of proteins in specific disorders via large scale throughput. Proteomics has been defined as "knowledge of the structure, function, and expression of all proteins in the biochemical or biological contexts of all organisms"(6). The identified proteins may be used for early detection of disease and also for a new therapeutic target (7).
The work of Hoang et al. (8) with 2DE and mass spectrometry might be the first report using the proteomic approach in placental trophoblasts. Hu et al. (9) presented 20 differential proteomic analysis of BeWo cell line in hypoxic conditions with 2DE and matrix-assisted laser desorption ionization time of flight mass spectrometry (MALDI-TOF-MS). As hypoxia is one of the initial events for development of preeclampsia the changes of proteomes are useful for the better understanding of preeclampsia. The relation with increased oxidative stress and reduced antioxidant defense, that suggested a pathogenic pathway, was demonstrated with preeclampsia-specific purified cytotrophoblasts by Johnstone et al. (10). More studies for the analysis of comparative proteomics using preeclampsia-specific purified cytotrophoblasts or mitochondria were reported by Sun et al. (11) and Shi et al. (12). In some research, differentially expressed proteins were also revealed in the placenta sample (13, 14, 15, 16, 17).
For these reasons, in this study we attempted to investigate the differences of protein expressions in the placenta sample between the preeclampsia and normal pregnancies for the understanding of pathogenesis and the discovery of helpful biomarkers.
MATERIALS AND METHODS
Subjects and sample collection
This study was designed for a case-control study. The placental tissue samples were obtained from five healthy pregnant women (the control group) and five preeclamptic pregnant women (the study group). To lessen the effects of labor and inflammation, all the pregnant women gave birth by cesarean section (breech presentation or repeated cesarean section) in the Obstetrics and Gynecology Department at Ajou University Medical Center. The control group was matched by maternal age, parity and gestational age at delivery with the study group and pregnant women with no history of other medico-surgical illnesses or obstetric complications were selected. In both groups, women with maternal infections, having fever and a maternal blood leukocytes count higher than 15,000/µL were excluded. Preecalmpsia was defined, when a pregnant woman with blood pressure showed 160/110 mmHg or higher blood pressure after the 20th week of gestation, measured at least twice with six hour intervals and if a urine analysis revealed 2 g or more of protein during a twenty four hour period and any findings of increased serum creatinine level (>1.2 mg/dL), decreased platelet count (<100,000 cells/µL), elevated hepatic enzyme activities (18).
Immediately following the cesarean section, the maternal side of the placenta and the central portion of chorion were dissected. The specimen were washed several times in 0.9% saline and then stored in a snap freezer with liquid nitrogen until protein extraction. Three spots were taken in each case.
Sample preparation, 2DE & image analysis
After grounding the 50-100 mg tissues were suspended in 4-5 volume sample buffer containing 7 M urea, 2 M thiourea, 100 mM DTT, 4% (w/v) CHAPS, 40 mM Tris, 0. 002% Bromophenol blue and protease inhibitor (Roche, Penzberg, Germany) and sonicated for protein extraction. Extracted protein concentration was determined with a Bradford assay kit (Bio-Rad, Hercules, CA, USA).
2DE was carried out essentially as described (19). One mg of total placenta protein was used for each electrophoresis. Aliquots in the buffer (7 M urea, 2 M thiourea, 4.5% CHAPS, 100 mM DTT, 40 mM Tris, pH 8.8) were applied to immobilized pH 3-10 nonlinear gradient strips (Amersham Biosciences, Uppsala, Sweden). Isoelectrofocusing was performed at 80,000 voltage hours. The second dimension was analyzed on a 9%-16% linear gradient polyacrylamide gel (18 cm×20 cm×1.5 mm) at a constant 40 mA per gel for approximately 5 hr. After protein fixation in 40% methanol and 5% phosphoric acid for 12 hr, the gels were stained with Coomassie brilliant blue G250 for 24 hr. The gels were de-stained with H2O, scanned in a Bio-Rad (Richmond, CA) G710 densitometer and converted into electronic files, which were then analyzed with an Image Master Platinum 5.0 image analysis program (Amersham Biosciences).
MALDI-TOF method
For the MALDI-TOF MS analysis, the peptides were concentrated by a POROS R2, Oligo R3 column (Applied Biosystems, Foster city, CA, USA). After washing the column with 70% acetonitrile, 100% acetonitrile and then 50 mM ammonium bicarbonate, samples were applied to the R2, R3 column and eluted with α-cyano-4-hydroxycinnamic acid (HCCA) (Sigma, St. Louis, MO) dissolved in 70% acetonitrile and 2% formic acid onto the MALDI plate (Opti-TOF™ 384-well Insert, Applied Biosystems) (20). MALDI-TOF MS was performed on a 4800 MALDI-TOF/TOF™ Analyzer (Applied Biosystems) equipped with a 355-nm Nd:YAG laser. The mass spectra were obtained in the reflectron mode with an accelerating voltage of 20 kV and sum from either 500 laser pulses calibrated using the 4,700 calibration mixture (Applied Biosystems).
Mascot database search
The ProFound (http://129.85.19.192/profound_bin/WebProFound.exe) was used to identify peptide sequences present in a protein sequence database. The database search criteria were, taxonomy, Homo sapiens (NCBInr database), fixed modification; carboxyamidomethylated (+57) at cysteine residues; variable modification; oxidized (+16) at methionine residues, the maximum amount of missed cleavage allowed, 1, MS tolerance, 100 ppm. Only peptides resulting from trypsin digests were considered.
Western blot analysis
After loading equal amounts of protein with 20 µg from each placenta and separation by 10% SDS-PAGE the proteins were transferred to Immobilion-P membranes, which were blocked overnight in Tris-buffered saline (20 mM Tris and 150 mM NaCl) and probed with primary antibodies of anti- heat shock protein (Hsp) 27 and anti- Hsp 70 (Santa Cruz Biotechnology, USA). The membranes were incubated with horseradish peroxidase-conjugated secondary antibody. As a control mouse monoclonal α-tubulin antibody was used. Densitometric analyses of the protein levels were measured. All experiments were performed three times per sample and showed similarities.
RESULTS
Characteristics of the study population
The maternal age, parity and gestational age at delivery did not show any significant differences because of matching at the time of enrollment. No significances were among the clinical parameters such as maternal platelet count and levels of serum creatinine. Systolic and diastolic blood pressure were significantly higher in the preeclampsia group than in the control group (181.3±6.5 vs. 117.8±5.1, P<0.0001; 112.0±13.2 vs. 73.8±9.8, P<0.0001, respectively). There were significant differences in the levels of hemoblobin and hematocrit between the preeclampsia and the control groups (13.7±1.2 vs. 10.9±0.8, P<0.05; 37.2±4.6 vs. 30.1±2.9, P<0.05, respectively). The levels of alanine aminotransferase and aspartate aminotransferase were significantly higher in the preeclampsia group than in the control group (194.8±37.6 vs. 24.0±10.6, P<0.05; 102.3±19.7 vs. 16.0±7.2, P<0.05, respectively). Neonatal birth weight was significantly lower in the preeclampsia group than the control group. The neonates with intrauterine growth resriction were excluded in the control group. The percentage of the baby with intrauterine growth retardation was 40% in preeclampsia (Table 1).
2DE expression and image analysis
The protein profiling of human placenta was performed using 2DE with five cases in each case of normal and preeclamptic pregnancy. The representative one in the preeclampsia and the control group are shown in Fig. 1. According to the percentage of intensity in each spot with comparative analysis, meaningful spots were determined using a statistical program.
MALDI-TOF MS data for identification of differentially expressed protein spots
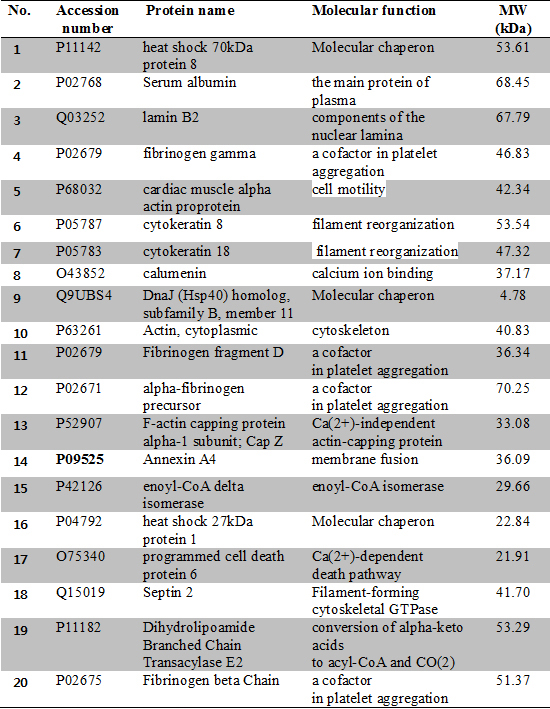
The protein maps showing more than twofold differences in each gel were marked as green colors in Fig. 2. Among these spots, twenty proteins that had significantly changed were selected (Table 2). The twenty spots from the 2D gels were picked up and identified using MALDI-TOF-MS (Fig. 3). The identified proteins in the MS data are listed in Table 3 in which there were 11 up-regulated proteins; fibrinogen gamma, cardiac muscle alpha actin proprotein, cytokeratin 8, calumenin, fibrinogen fragment D, F-actin capping protein alpha-1 subunit, Hsp 27, Hsp 40, annexin A4, enoyl-CoA delta isomerase and programmed cell death protein 6 and 9 down-regulated proteins; Hsp 70, serum albumin crystal structure chain A, lamin B2, cytokeratin 18, actin cytoplasmic, alpha fibrinogen precursor, septin 2, dihydrolipoamide branched chain transacylase E2 and firbrinogen beta chain (Table 3).
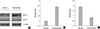
The expression of Hsp 27 and Hsp 70 protein by the Western blot analysis
To validate the expression of proteins in 2DE gels immunoblotting for Hsp 27 and Hsp 70 were performed. As shown in Fig. 4 the significant up-regulation of Hsp 27 and down-regulation of Hsp 70 were demonstrated by the Western blot analysis.
DISCUSSION
The proteomics technology has been used to specifically determine cellular or circulating protein expressions that may be involved in the pathogenesis of disease (16). The etiology of preeclampsia is explained by the placental ischemia caused by abnormal trophoblastic invasion to spiral arteries, immunologic intolerance and inflammatory change. As a consequence, inflammatory cells are released and they induce oxidative stress to cause endothelial dysfunction and endothelial cell injury. Also, recent studies suggest that apoptosis is closely related to preeclampsia and that placental ischemia contributes to increased apoptotic activity (5). For elucidation of this complexity our proteomic study might add the candidate proteins into the journey for an explanation of the preeclamptic pathogenesis. All twenty differentially expressed proteins- down regulated or up regulated-show an association with the suggested pathogenic mechanism of preeclampsia. One of the significant proteins identified in this study is the heat shock protein acting as a molecular chaperon in the surviving cells under stressful conditions (21). Among these the class of molecular chaperones Hsp 70, 40 and 27 were identified in this study. Hsp70 is also a stress-induced protein and this protein can be an anti-apoptotic protein and an oxidative stress marker for ischemic and apoptotic circumstances like preeclampsia. These proteins show not only multifunctional molecular chaperones which are involved in protein folding, degradation, and transport but also roles in the suppression of anti-inflammatory cytokines (22, 23). In preeclampsia, the change is contradictory. The increased placental levels of Hsp70 have been reported in the proteomic approach (24, 25, 26). The reduced results that are similar to this study have also been presented by Johnstone et al. (10). Further study for the reasons behind reduced levels being either one of the original pathogenic factors or secondary effects in preeclamptic placenta should be evaluated.
The increased expression of Hsp 27 is particularly interesting as it can primarily induce anti-apoptotic responses. Our data support the up-regulation of Hsp 27 in several other studies (13, 14, 27). In addition to increased Hsp 27, the increased levels of phosphorylated Hsp 27 were shown in preeclampsia by the proteomic technique followed by verification with the Western blot analysis (14).
The discovery of up regulation in Hsp 40, using proteomics in a human placenta is the first time it has been reported to my knowledge. As the activity of Hsp 70 is determined by Hsp 40 the interaction between two proteins seems to do very important roles in the progression of preeclampsia. Despite several studies demonstrating the role of Hsps in preeclampsia it is still under debate and more studies are essential.
Beta oxidation is the process that fatty acid molecules are broken down in the mitochondria to generate acetyl-CoA used by the electron transport chain (28). Enoyl-CoA isomerase is known as dodecenoyl-CoA isomerase or acetylene-allene isomerase. It has a particularly important role in the metabolism of unsaturated fatty acids. The altered fatty acid oxidation in a preeclamptic placenta has been speculated as one of the pathophysiology of preeclampsia. Interestingly, up regulation of enoyl-CoA isomerase showed in this study contrary to the down regulation that is shown in other reports with preeclampsia placentas using the proteomics method (29, 30). Therefore the altered regulation of involved enzymes in beta oxidation - down regulated or up regulated- can induce other changes in preeclampsia. Shi et al. (12) also proposed that fatty acid oxidation is one of the critical processes in the development of preeclampsia using quantitative mitochondrial proteomics. Researching further will contribute to the understanding of the pathophysiology of preeclampsia through the pathway analysis.
We found a lower expression of dihydrolipoamide branched chain transacylase E2, an inner-mitochondrial enzyme complex involved in the breakdown of the branched-chain amino acids isoleucine, leucine, and valine. This has not been reported to be expressed in preeclamptic placenta. The pathologic role of the down regulation should be cleared with regard to mitochondria dysfunction.
The expression of structural proteins for cytoskeleton components, which are cytokeratin 8 and cytokeratin 18, were found to be up-or down-regulated. These proteins have been implicated for cell mobility and cytoskeletal structure formation. Hu et al. (9) presented similar results like this study.
Annexins are a family of calcium- and phospholipid-binding proteins which are involved in diverse cellular functions, membrane fusion, signal transduction, apoptosis, growth regulation and inflammation links with various human diseases like cancer, cardiovascular disease, and brain ischemia (31). The Annexin A4 is one of the annexin family that has anticoagulant activity, inhibition of phospholipase A2 activity and apoptosis was expressed with up regulation in this study (32). It has been generally shown that under oxidative stress conditions, increased phospholipase A2 activity would lead to an increase in decomposition of arachidonic acid, finally resulting in more production of thromboxane A2. On the other hand, the over-expression of annexin A4 in rat glioma C6 cells enhanced ethanol-induced cell lesions, accompanied by nuclear factor-kappaB (NF-κB) activation (33). NF-κB is a transcription factor which plays an important role in the cell proliferation, inflammation, invasion, angiogenesis, apoptosis, and cellular adhesion that can be activated by oxidative stress, proinflammatory cytokines, growth factors, death receptors, and DNA-damaging agents, which are important features in suggested pathogenesis in preeclampsia (34). Also, Jeon et al. (35) clearly demonstrated that annexin A4 differentially modulates the NF-κB signaling pathway via interactions with the p50 subunit of NF-κB in a Ca2+-dependent manner. To our knowledge, it is the first research to show the relationship between significantly up-regulated annexin A4 and preeclampsia and an involved mechanism should be revealed through further detailed study. The reason for the up-regulation of annexin A4 might be compensated by antioxidant activity or not. This would be determined through further analysis.
Other members of the annexin protein family, annexin A2 and A5 were observed with increased expression under hypoxia for the processs of trophoblast syncytialization by Hu et al. (9) and annexin A1, A6, and A11 appeared with up regulation by Wang et al. (15).
The work of Wang et al. (15) detected a total of 2,636 proteins in the human placenta and 171 differently expressed proteins between preeclamptic placentas and healthy pregnant placentas. The identification of significantly different expression proteins are much more numerous compared to other studies, 17 proteins in Kim et al. (13), 20 proteins in Hu et al. (9) similar with this study, 12 proteins in Shin et al. (14), and 11 proteins in Johnstone et al. (10). Even with the development of technology as time goes by and different samples such as total placentas or cultured trophoblasts, the big difference between Wang et al. (15) and other researches including this report, presumed the use of the LC technique and 2DE. Although the number of total identified proteins is smaller, 2DE can identify more structural proteins like cytoskeleton components and proteins associated with coagulation pathway or platelet aggregation than the LC technique. The programmed cell death protein 6 identified in our work is the only protein consistent with the findings of Wang et al. (15). Despite the fact that 2DE and the LC technique can detect a different spectrum of proteins, because the coverage fields of identified proteins have differences, both methods collectively contribute to give valuable information to understand the involved mechanism of preeclampsia. Another difference between our study and that of Wang et al. (15) is the severity of preeclampsia. The more severe clinical manifestations including higher systolic and diastolic blood pressure; elevated liver enzymes, and abundant proteinuria, these can result in different comparative proteome profiles according to the severity of the disease. The confirmation of suggestive pathogenesis of preeclampsia including apoptosis, oxidative stress, and fatty acid oxidation, enrollment of cases with severe clinical conditions, and detection of novel proteins as possible biomarkers are advantages in this study.
In conclusion, our proteomic findings in preeclamptic placentas showed consistent findings with defective anti-oxidative stress response, anti- inflammation and anti-apoptosis including changes in several proteins participating in these processes. These proteins are programmed cell death protein 6, annexin A4, enoyl-CoA isomerase, dihydrolipoamide branched chain transacylase E2, Hsp27 and Hsp70 in support with suggested pathogenesis of preeclampsia (36). The progression of cumulative data previously performed in proteomics research and newly identified proteins included in our study, make contributions to understand the pathogenesis of preeclampsia through the systematic investigation in changes of critical proteins.





 XML Download
XML Download