

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Prostate cancer is the second most frequently diagnosed cancer affecting male adults and the sixth most common cause of cancer associated death in men across the world (1). The major risk factors for prostate cancer are old age, race, inherited susceptibility and environmental and behavioral factors such as diets (2, 3). Treatment options of prostate cancer include active surveillance, surgical resection, androgen ablation therapy, radiotherapy, and cryotherapy variously depending on risk grouping, medical comorbidities and age at the diagnosis of the patients (4, 5, 6).
Prostate cancer is heterogeneous in terms of clinical behaviors, histological and molecular features, and treatment options (4, 5, 6, 7). Although most patients with low and intermediate-risk prostate cancer follow an indolent clinical course and are potentially curative by surgical resection or radiotherapy, patients with high-risk prostate cancer have higher rate of cancer related death and conventional chemotherapy and radiation therapy are of limited effectiveness for them (2, 4, 5, 6, 8). Androgen deprivation therapy has good efficacy for advanced disease to lead to either a partial or complete remission initially. However most patients evolve towards an androgen-independent state within a few years and result in death due to widespread metastasis (6). Therefore, more improved treatments and novel biomarkers are strongly needed in order to overcome metastasis and lethal recurrence for advanced prostate cancer as well as unpredictable cases of low or intermediate risk groups.
Peroxisome proliferator-activated receptors (PPARs) are a group of 50-kDa ligand-dependent transcription factors localized at human chromosome 3p25 and belong to a member of the nuclear hormone receptor superfamily such as steroid and thyroid hormones (9, 10, 11). They are composed of three isoforms (PPAR-α, PPAR-γ, and PPAR-δ/β) and require heterodimerization with the retinoid X receptor for optimal DNA binding to specific response elements, termed to as peroxisome proliferator response elements (PPREs), in the promoter of the various target genes (9, 10, 11). The nuclear receptor superfamily was named because they show prominent nuclear localization.
Among the subtypes, PPAR-γ is known to play a prominent role in adipocyte differentiation, the inflammatory response and peripheral glucose utilization (9, 11). Recent studies have suggested that PPAR-γ play a key role in tumorigenesis as a tumor suppressor and PPAR-γ agonists showed antiproliferative and proapoptotic actions in cancers (8, 12, 13). Currently PPAR-γ agonists including the most active natural ligand, 15-deoxy-D-12, 14-prostaglandin J2 (15-dPGJ2) and the most specific PPAR-γ synthetic agonists, the thiazolidinediones (TZDs) are used for enhancing insulin sensitivity in type II diabetes patients (9). And they have been introduced in clinical trials for treatment of several malignancies such as colorectal and esophageal cancers (9, 14).
Many in vivo or in vitro studies have shown antineoplastic effect of PPAR-γ ligands in prostate cancers (15, 16). It has been reported that exposure to the TZDs has been shown to increase apoptotic activity in LNCaP, C4-2, and PC-3 prostate cancer cells and a combination therapy using histone deacetylases inhibitors and PPAR-γ agonists decrease invasiveness of the prostate adenocarcinoma cells in vivo (13, 15, 16). Efatutazone is recently introduced as a novel third-generation TZD PPAR-γ agonist, which is at least 500-fold more potent than the other TZDs such as troglitazone and pioglitazone (17). Therefore, PPAR-γ and its agonists have recently drawn more attention and promise in terms of chemoprevention and chemotherapy for cancer treatment and it has been also considered as a promising molecular target for anticancer targeted therapy development in prostate cancers (18).
However, there is little or controversial information on PPAR-γ expression and clinical implications in the clinical specimen of prostate cancers (18, 19, 20). Whereas Nakamura et al. (19) reported PPAR-γ expression showed an inverse correlation with worse clinical parameters including pT stage and serum PSA levels, Rogenhofer et al. (20) reported PPAR-γ protein and mRNA expression were found to be significantly higher in advanced prostate cancers than in localized cancer. Therefore, further studies are needed to obtain a better understanding of the clinical roles of PPAR-γ in human prostate cancers for considering a potential trial of PPAR-γ ligands therapy for the patients.
The aim of this study was to validate PPAR-γ expression in malignant and benign prostate tissues of a clinically well-characterized prostatectomy cohort by immunohistochemistry and quantitative real-time PCR in a Korean population.
MATERIALS AND METHODS
Case selection
A retrospective study was conducted in 730 Korean patients who underwent radical prostatectomy and were subsequently diagnosed as prostatic adenocarcinoma (PCA) between 2005 and 2012 at the Departments of Pathology, Konkuk Medical Center and Asan Medical Center, Seoul, Korea. All histological sections were reviewed and appropriated representative sections for each study were selected. Tissue microarrays (TMAs) including 667 PCAs were made to investigate PPAR-γ expression in PCAs by immunohistochemistry and subsequently the representative whole section of 63 radical prostatectomy specimens including PCA and benign prostate tissue were included to validate the results of TMA study. Clinicopathologic data including age, Gleason score and pathologic stage were collected from pathology reports. Pathologic T stage was assigned according to the 7th edition American Joint Committee on Cancer TNM system (21).
Construction of tissue microarray
TMAs were constructed using a manual tissue arrayer (Beecher Instruments, Sun Prairie, WI, USA). Three randomly representative 0.6 mm cores were obtained from the most representative caner areas of formalin-fixed paraffin-embedded tissue blocks and were arranged in TMA blocks.
Immunohistochemistry
The representative paraffin blocks from 63 radical prostatectomy specimens and TMAs including 667 PCAs were sectioned into 4 µm slices, deparaffinized, and antigens demasked in EDTA buffer, pH 8.5 (Cell Conditioning 1 solution, Ventana). Immunohistochemical analysis was conducted with the Ventana Discovery XT automated staining system (Ventana Medical Systems, Tucson, AZ, USA) according to manufacturer's instruction. Staining of two PPAR-γ antibodies (C26H12, rabbit monoclonal, 1:400, Cell Signaling Technology, Danvers, MA, USA; H100, rabbit polyclonal, 1:25, Santa Cruz Biotechnology, Santa Cruz, CA, USA) was used according to each manufacturer's instruction. Detection of PPAR-γ was carried using the iVIEW DAB Detection Kit (Ventana). All staining reactions were carried out in Tris buffer, pH 7.4-7.8 (Ventana reaction buffer, Ventana). Finally, slides were counterstained with hematoxylin I, followed by bluing reagent (Ventana). Omitting of primary antibody was used for the negative control and urothelial carcinoma in bladder and thyroidal follicular carcinoma were used for the positive control.
RNA extraction
The H&E slides from all samples were reviewed by two pathologists to mark the areas with the collected sections of 63 prostatectomy specimens composed of 10 PCAs with low Gleason score (≤7, Low GS), 10 PCAs with high Gleason score (>7, High GS), five benign prostate tissues as a control for fold change, and thyroidal follicular carcinoma showing PPAR-γ nuclear staining confirmed by immunohistochemistry as positive control. Subsequently, 10 µm thick sections cut from the formalin-fixed, paraffin-embedded blocks for dissecting the target area from the slides, then, transferred into microcentrifuge tubes. Before RNA isolations, deparaffinization was performed with 1 mL xylene with 10 min incubation time, and 1 mL absolute ethanol for 10 min. Total RNA was extracted by using High Pure FFPE RNA Isolation Kit (Roche Applied Science, Penzberg, Germany), according to the manufacturers' instructions. RNA yield and quality was assessed by measuring the ratio of spectrophotometric absorbance (260 nm/230 nm and 260 nm/280 nm) using NanoDrop® ND-1000 Spectrophotometer (Thermo Fisher Scientific Inc., Waltham, MA, USA). The extracted RNA samples were considered to be acceptable only with absorbance ratio between 1.0 and 2.0 at 260 nm/230 nm, and between 1.8 and 2.0 at 260 nm/280 nm.
Quantitative real-time polymerase chain reaction
Complimentary DNA was synthesized by reverse transcription using Transcriptor First Strand cDNA Synthesis Kit (Roche Diagnostics, Mannheim, Germany). 1 µg of total RNA were reverse transcribed with a combination of anchored-oligo (dT) and random hexamer primers, according to the manufacturers' instructions. To assess PPAR-γ (Hs01115513) and GAPDH (Hs99999905) gene expression, we used pre-designed quantitative real-time TagMan Gene Expression Assays (Applied Biosystems, Foster city, CA, USA). Quantitative real-time PCR reactions were performed in final volume of 20 µL using 10 ng cDNA/well and 10 µL FastStart Essential DNA Probes Master (Roche Diagnostics) according to the manufacturers' instructions. Thermal cycling conditions on the LightCycler Nano System were the following: Enzyme activation: 95℃ for 10 min, 45 cycles of amplification: 95℃ for 10 sec, 60℃ for 30 sec. Each quantitative real-time PCR analysis was performed in triplicate. The glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as the internal control gene. The normalized amount of each target mRNA present in 20 PCAs cases was calculated by benign prostate tissue cases.
Statistical analyses
The unpaired t-test, chi-square test or Linear-by-Linear Association test were used to compare the PPAR-γ immunoreactivity in accordance with the clinicopathologic variables including age, pT stage and Gleason score. The unpaired t-test was also used to compare the relative mRNA expression level between groups and controls. P<0.05 was considered significant. All statistical analyses were conducted using SPSS v. 17.0 (SPSS, Inc., Chicago, IL, USA).
RESULTS
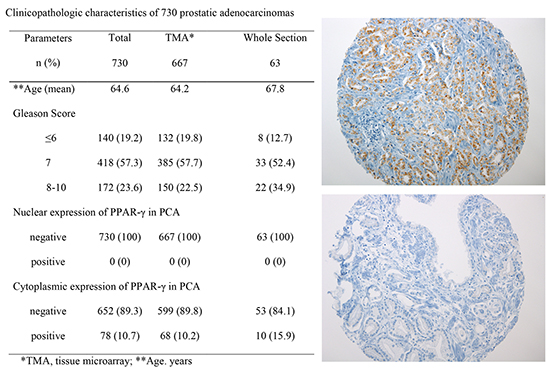
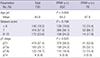
The clinicopathologic features of the PCAs from 730 patients including age, Gleason score and pathologic T stage enrolled in this study are summarized in Table 1. Patient ages ranged from 42 yr to 89 yr (mean, 64.6 yr). Gleason score were lower than 7 in 19.2% of the patients (140/730), 7 in 57.3% of the patients (418/730), and higher than 7 in 23.6% of the patients (172/730). 57.4% of the patients were in pT2 stage (419/730), 25.1% in pT3a stage (183/730), 16.7% in pT3b stage (122/730), and 0.8% in pT4 stage (6/730) (Table 1).
The subcellular localization and expression of the PPAR-γ protein were evaluated on TMAs including 667 PCAs by immunohistochemistry. Subsequently 63 whole sections of radical prostatectomy with PCA and adjacent benign prostate tissue were incorporated in this study to validate TMA's results. None of 730 PCAs showed nuclear immunoreactivity of PPAR-γ (Fig. 1A) (Table 1). 10.7% of PCAs (78/730) showed diffuse cytoplasmic immunoreactivity of PPAR-γ; 10.2% of PCAs (68/667) from TMA (Fig. 1B) and 15.9% of PCAs (10/63) from the whole sections (Fig. 2A-C) (Table 1). Tumor heterogeneity of PPAR-γ immunoreactivity was identified in PCAs (Fig. 2D).
Most benign prostatic glands showed negative immunoreactivity of PPAR-γ except for variable weak cytoplasmic staining of basal cells in some glandular epithelum (Fig. 3A). Verumontanum mucosal epithelium showed nuclear staining of PPAR-γ (Fig. 3B) and some glandular epithelium of benign prostatic hyperplasia revealed nuclear and/or cytoplasmic immunoreactivity of PPAR-γ (Fig. 3C and D).
Positive controls revealed nuclear immunoreactivity of PPAR-γ in thyroid follicular carcinoma, urothelial carcinoma of bladder and periprostatic adipocytes (Fig. 4A-C). No association between PPAR-γ cytoplasmic expression and clinicopathologic parameters including age, Gleason score and pathologic stage was noted (Table 2).
The expression level of PPAR-γ mRNA for each sample was standardized based on the mean expression level of five benign prostate tissues as a control for fold change. The mean fold change indicated that the constitutive PPAR-γ mRNA in PCAs (mean fold change=0.68) was marginally detectable compared to that of thyroid follicular carcinoma as a positive control (mean fold change=18.11) (Fig. 5A). The expression of PPAR-γ mRNA were down-regulated in low GS (mean fold change 0.67, P=0.007) and high GS (mean fold change 0.66, P=0.022) groups when compared to that of the benign prostate tissues (Fig. 5A).
10% of low GS (1/10) and 33.3% of high GSs (3/10) showed higher expression of PPAR-γ mRNA compared to that of the benign prostate tissues (Fig. 5B). There were no significant difference of mRNA expression of PPAR-γ between the low GS and high GS groups (P=0.905) (Fig. 5B). There is no significant difference of mRNA expression between cytoplasmic immunopositive PCAs and immunonegative PCAs of PPAR-γ (P=0.88).
DISCUSSION
PPAR-γ expression has been investigated in various human diseases including cancers, diabetes, atherosclerosis, and neurodegenerative disease (9, 11, 22). The PPAR-γ gene is known to function as a tumor suppressor gene involved in tumor growth inhibition, apoptosis and tumor cell differentiation (9, 15, 17). It has been mapped to chromosome 3p25, of which frequent heterozygous deletions are observed in human cancers including prostate cancer (9). Its expression level is known to be varied dependent on different types of tissues and carcinomas (23). This receptor has initially shown nuclear localization of genomic activity via direct or indirect transcription as a key regulator of metabolic homeostasis and inflammatory response (10, 11, 18). Recently the nongenomic activity and the cytoplasmic localization of the receptor have been also magnified as new challenging from a functional perspective in the diseases (10, 24, 25).
Our results revealed that PPAR-γ was marginally detectable in prostate tissues of Korean population and the localization of the receptor was mainly in the cytoplasm, not in the nucleus. We first planned this study under the hypothesis that PPAR-γ would be expressed in the nuclei of tumor cells as reported in previous studies that showed its nuclear immunoreactivity in over 70% of prostate cancer (19, 20). These conflicting findings prompted us to discuss several questions from all angles including extranuclear localization of PPAR-γ, ethnical variability and technical issues of immunohistochemistry.
PPAR-γ is known to exert pleiotropic effects in glucose and lipid metabolism, anti-inflammatory responses, apoptosis and carcinogenesis by modulating specific gene expression (9, 11, 26). A number of studies have described that the PPAR-γ localize predominantly in the nucleus to exert its genomic effect via direct or indirect enhancement or inhibition of transcription (9, 11, 26). However, recently new convincing evidences of the nongenomic activity of the PPAR-γ and their cytoplasmic localization have been reported in many studies (10, 16, 18, 24, 25). Burgermeister and Seger described that PPAR-γ has cytoplasmic activities that are caused by mitogenic stimulation suppressing PPAR-γ's genomic activity and MEK-dependent shuttle may also play a role in the control of the extra-nuclear/nongenomic actions of PPAR-γ (25). In this study, the cytoplasmic localization of PPAR-γ would be also noticeble to support extra-nuclear/nongenomic actions of the PPAR-γ.
Recently, Lee et al. demonstrated PPAR-γ was localized predominantly in the perinuclear region and cytoplasm in the untreated prostate PC-3 and LNCap cells and 4-O-methylhonokiol (MH), a constituent of Magnolia officinalis increased the protein and transcriptional expressions of PPAR-γ via its nuclear translocation from the cytoplasmic location in those cells (16). As most previous studies has demonstrated mainly quantitative or qualitative analysis of PPAR-γ, Lee et al.'s study showing remarkable morphologic localization of PPAR-γ protein in prostate cancer cell lines is noteworthy. It corresponded with its cytoplasmic location in clinical specimen with prostate cancer in the current study. It is also very interesting to draw a hypothetical reasoning that PPAR-γ can play a significant and different role as a nuclear transcription factor in in vivo human prostate cancers when it is induced from cytoplasm to nuclei by its agonists.
Although many studies have indicated beneficial effects with PPAR-γ ligand treatment, the role of PPAR-γ in cancer therapy is still controversial showing either pro-tumorigenic or antineoplastic effects in studies (12, 13, 27). Nonetheless, many previous studies have shown that TZDs decrease the growth rate of the human prostate cancer cells both in vitro and in vivo (13, 28). And two clinical trials using the TZD troglitazone showed effective therapeutic results in the patients with prostate cancer (9, 29). Recently Sawayama et al. demonstrated efatutazone, a novel third-generation TZD PPAR-γ agonist which is at least 500-fold more potent than troglitazone alone or in combination with cetuximab may offer as a potential therapy for esophageal squamous cell carcinomas (17). They also demonstrated efatutazone combined with cetuximab showed synergistic antiproliferative effects by suppressing both the PI3K-Akt and MAPK pathways. Therefore, it will be important in the further experiment with these promising TZD drugs which may improve the prostate cancer prognosis.
Our study demonstrated PPAR-γ protein and mRNA expression are low in nonneoplastic and neoplastic prostate tissues of Korean population, and PPAR-γ mRNA expression was observed to be slightly lower in prostate cancers than in the benign prostatic glands. These results were supported by several articles that PPAR-γ mRNA expression in carcinomas was lower than that in normal prostatic tissues (12, 23). However, the up-regulation of PPAR-γ expression is a frequent occurrence in a variety of different malignant tumors (30, 31). Segawa et al. reported high expression level of PPAR-γ mRNA in prostatic intraepithelial neoplasia (PIN) and prostatic carcinoma compared to low expression level of PPAR-γ mRNA in benign prostatic hyperplasia, and no expression of PPAR-γ in normal prostatic tissue (32). Our study has some limitations because of small sample size, fixed tissue for RT-PCR and lack of in situ hybridization. On the other hand, as PPAR-γ activity is known to be modulated directly and indirectly particularly at its transcriptional activity regulated by post-translational modifications (10, 25), further research will be necessary to clarify these controversial findings.
These controversies in PPAR-γ expression could be elucidated that the pathogenesis of prostate cancer reflecting both hereditary and environmental components is ethnically different (7, 26). The genetic changes as well as the incidence and prognosis of prostate cancer have shown strong ethnic variance, particularly in Asian populations (7). For example the frequency of transcription regulator ERG gene is much lower in Asian countries in contrast to higher ERG frequency in Western countries (33, 34). As a possible association between PPAR-γ polymorphism and the risk of developing cancer has been reported (26, 35), polymorphisms in the PPAR-γ gene that show increased or decreased function variants may cause the different expression and role of PPAR-γ in prostate cancers of Korean men. Epidemiological studies have shown certain environmental elements such as the consumption of red meat and animal fats have had an effect on the increased rate of prostate cancer (3). In Korea, prostate cancer is the fifth most frequently diagnosed cancer in males and has been rapidly increased according to transition to Western dietary habits among Korean (36). These perspectives may implicate a link between PPAR-γ's function involved in lipid metabolism and its role in prostatic carcinogenesis.
On the other hand, in recent oncologic research and medicine, accuracy of immunohistochemistry to validate the target genes is very important step to move to the further effective target therapy (37). We used three appropriate positive controls for PPAR-γ immunostaining, which demonstrated the positive nuclear immunoreactivity well in urothelial carcinoma and follicular carcinoma as well as in periprostatic adipocytes as the internal control. We also tested primary antibody using two different PPAR-γ antibodies that are widely used in the literatures, which confirmed the same results by both antibodies at two different hospital laboratories. The previous literature have described both nuclear and cytoplasmic immunoreactivity of PPAR-γ, but revealed much nonspecific background staining of PPAR-γ immunoreactivity in the given photos (18, 19) with no positive or negative control information. Commercially available and well characterized PPAR-γ antibodies have been recently introduced to clinical and research communities, therefore, further study is needed to investigate the expression pattern of PPAR-γ using an optimized immunohistochemistry in a large cohort of clinical specimens with prostate cancers.
In conclusion, our study demonstrated the cytoplasmic localization of PPAR-γ protein expression supporting its nongenomic cytoplasmic activity in carcinogenesis of prostate cancer. Further investigations are necessary to clarify clinical or biological roles of PPAR-γ and to provide useful information of PPAR-γ agonists for a new therapeutic option in the patients with prostate cancers.



 XML Download
XML Download