

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Lumbar disc herniation can injure spinal nerve roots and cause severe radicular pain, characterized by hyperalgesia and allodynia. Previous studies have suggested the symptoms of lumbar disc herniation are caused by chemical factors released from the nucleus pulposus and by mechanical compression of the lumbar nerve root (123) and that they are mediated by proinflammatory cytokines derived from the herniated intervertebral disc (45). Nucleus pulposus contains tumor necrosis factor alpha (TNFα), and nucleus pulposus applied to dorsal root ganglions (DRGs) induces pain-related behavior in rats and morphological and functional changes in DRGs and spinal cords (1234). Several studies have reported increases in prostaglandins, pain-associated neuropeptides (e.g., substance P [SP], calcitonin gene-related peptide [CGRP]), cytokines (e.g., TNFα), and ion channels in DRG neurons and the spinal dorsal horns in animal models of disc herniation (67).
Clinically, lumbar disc herniation is usually diagnosed based on symptoms, signs, and imaging findings, particularly magnetic resonance imaging (MRI) findings, as these provide strong diagnostic evidence following careful history taking and physical examination. However, the syndrome of lumbar radiculopathy often affects areas far beyond the innervation areas of affected nerves and is not consistent with MRI findings, which makes identification of the location of the disease and treatment difficult and may even lead to mismanagement. Furthermore, the explanation of this phenomenon remains unclear.
Studies have been conducted on the effect of lumbar disc herniation on neurotransmitter dynamics in primary sensory neurons and primary afferent nerve fibers (89). Badalamente et al. (8) confirmed that mechanical compression of DRGs in rats resulted in SP concentration increases in ipsilateral DRGs cells and in spinal dorsal horns, as determined by immunofluorescence and using isotopes. Kobayashi et al. (9) found compressive disturbance of nerve roots caused Wallerian degeneration at the site of nerve root compression and at synapses of spinal cord dorsal horns. These studies suggest disruption of axonal flow in the central branches of the primary sensory fibers caused by disc herniation disrupts neurotransmitter metabolism in synapses of the spinal dorsal horn and in sensory neurons of DRGs and that these disruptions are closely related in the onset of pain and sensory disturbances. Thus, it appears that various neurotransmitters produced by primary sensory neurons in the DRG and transported to the spinal dorsal horn by axonal flow in the central branches of dorsal root neurons are implicated in the development of radicular pain and sensory disturbance associated with nerve root compression.
However, knowledge and mechanistic evidence about changes in adjacent segments of dorsal horn and DRG are lacking. We hypothesized that neural pathways exists between adjacent lumbar spinal cord segments and dorsal root ganglia. Accordingly, we examined the expressions of ionized calcium-binding adapter molecule 1 (Iba1), glial fibrillary acidic protein (GFAP), CGRP, and transient receptor potential vanilloid 1 (TRPV1) in ipsilateral adjacent segments in a rat model of radiculopathic pain.
MATERIALS AND METHODS
Animals
Twenty-nine male Sprague-Dawley rats (200-250 g) were randomly assigned to either a sham-operated group (n=10) or a nucleus pulposus (NP)-exposed group (n=19). Rats were housed two per cage and had free access to water and food.
Lumbar disc herniation
Rats were anesthetized by intraperitoneal injection of Zoletil (Virbac) at 50 mg/kg. With an animal placed prone, an incision of approximately 1 cm was made on the dorsal surface of the proximal tail for autologous nucleus pulposus harvesting. The disc between the second and third coccygeal vertebrae of each tail was incised, and nucleus pulposus was harvested by curette. A midline dorsal incision was then made over the lumbar spine, multifidus muscles were separated along L4-S1 spinous processes, and left L5 nerve roots and DRG were exposed by laminectomy. The harvested nucleus pulposus was then implanted next to the left L5 nerve root just proximal to the DRG without mechanical compression. Similar amounts of nucleus pulposus were implanted in all animals. The sham procedure was performed in an identical manner and included autologous nucleus pulposus harvesting and nerve root exposure but not autologous nucleus pulposus implantation (1011).
Pain behavior evaluation
Mechanical sensitivity of the plantar surfaces of ipsilateral hind paws was tested at 10 days after surgery. Mechanical allodynia was assessed by measuring withdrawal responses of ipsilateral hind paws to mechanical stimulation with von Frey filaments (North Coast Medical, Inc., Gilroy, CA, USA), which were calibrated in grams. Briefly, a rat was placed in a clear plastic cage with a metal mesh floor, allowed to adapt to the testing environment for 30 min, and then the plantar surface of each hind paw was stimulated so as to cause slight filament bending for 5 sec. Filaments were applied in increasing and decreasing thicknesses, beginning with a 0.1-g probe, until a filament produced a consistent withdrawal response to more than 3 of 5 stimuli. We calculated 50% probability thresholds of mechanical paw withdrawal. If no withdrawal response was elicited by the 26-g (the thickest) filament, the mechanical threshold was assigned as 26 g. Rats in the NP group were divided into 2 subgroups according to measured mechanical thresholds. Animals with a mechanical threshold of <5 g were assigned to significant pain (n=12) or ≥5 g were no significant pain subgroups (n=7) (12).
Immunohistochemical examination
To study microglial and astrocytic activation in the dorsal horn and CGRP and TRPV1 expressions in DRGs, we euthanized all 29 rats at 10 days after disc herniation surgery. Under anesthesia, a catheter was inserted into the left ventricle, which was then rinsed with 500 mL of saline and then fixed with 500 mL of 4% paraformaldehyde (in 0.1 N phosphate buffer [PB]).
Spinal cords at the L3-S1 level were removed, post-fixed for 2 days in the same fixative, and stored in 30% sucrose (in PB) for at least 24 hr. Transverse sections (30 µm) of each spinal cords (from corresponding spinal nerve root to inferior border of upper spinal nerve root) (Fig. 1) and of DRGs (20 µm) were prepared using a cryostat (Leica, Wetzlar, Germany) and stored in PB. All incubation and reaction procedures for multiple immunohistochemical staining were performed at room temperature on a shaker. To enhance antibody penetration into tissues, DRG sections were immersed in 50% ethanol for 30 min and rinsed with phosphate buffered saline (PBS) for 5 min, three times, and to block nonspecific primary antibody reactions, samples were treated with 10% normal donkey serum (NDS; Jackson Immunoresearch, Westgrove, PA, USA). Tissue sections were incubated overnight in a mixture of the following primary antibodies; mouse anti-Ionized calcium-binding adapter molecule 1 (Iba1) (Wako, Osaka, Japan; 1:1,000), mouse anti-glial fibrillary acidic protein (GFAP) (BD Pharmingen, San Jose, CA, USA; 1:100), anti-transient receptor potential vanilloid type 1 (TRPV1) (Neuromics, Edina, MN, USA; 1:5,000) and anti-calcitonin gene related peptide (CGRP) (Enzo, Farmingdale, NY, USA; 1:200). Tissues were then rinsed with PBS for 5 min, three times, treated with 2% NDS for 15 min, and incubated with cy3-conjugated donkey anti-mouse (Jackson Immunoresearch, PA, USA, 1:100), cy3-conjugated donkey anti-goat (Jackson Immunoresearch, PA, USA, 1:100) and Alexa 488-conjugated donkey anti-rabbit (Invitrogen, Eugene, OR, USA, 1:200) antibodies for 3 hr, and then rinsed with PBS and mounted with Vectashield (Vector Lab, Burlingame, CA, USA). All antibodies were tested for sensitivity and specificity before the study, and manufacturers' recommended dilutions were used. Immunofluorescent images were acquired using a cooled charge-coupled device (CCD) camera (Olympus DP71, Tokyo, Japan) attached to a light microscope (Olympus BX51).
Quantitative image analysis
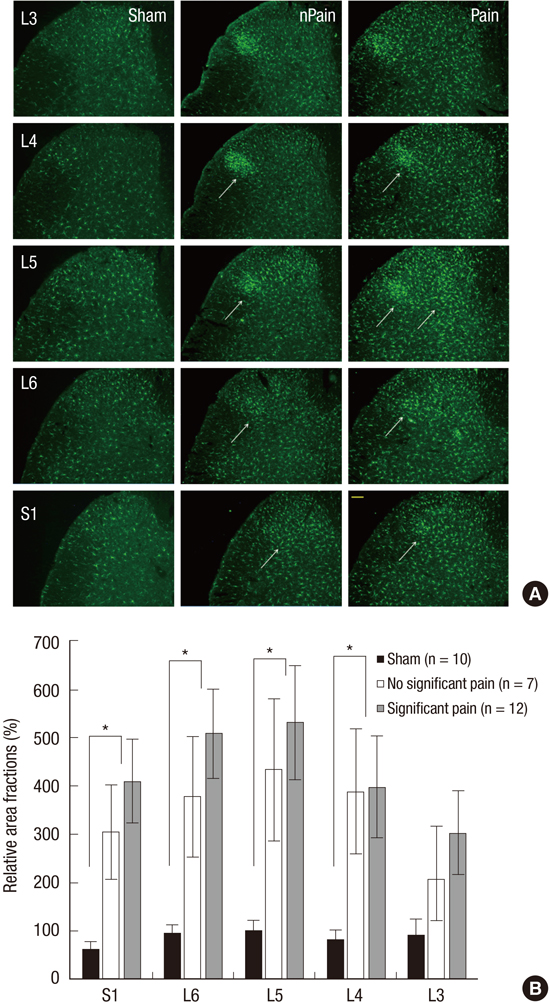
To analyze immunoreactions of Iba1 and GFAP in dorsal horns and of CGRP and TRPV1 in DRGs quantitatively, we obtained images from five spinal cord sections (for Iba1 and GFAP) from L3, L4, L5, L6, and S1 segments and 5 DRG sections from L3, L4, L5, L6, and S1 DRGs (for CGRP and TRPV1) per rat. One image (898×660 mm) was acquired of each spinal cord section using a CCD camera using the same shutter speed and digital gain. Images were encoded in order to blind the investigator before analysis. Pixels positive for Iba1 and GFAP immunoreactions were segmented by applying an appropriate threshold gray value and area fractions (segmented area/total frame area) were calculated using image analysis software (Leica application suite V4.2, Leica Microsystems, Heerbrugg, Switzerland). In case of CGRP and TRPV1, numbers of CGRP- and TRPV1-postive DRG cells were counted in the same manner using image analysis software. Then, relative area fractions of Iba1 and GFAP immunoreactions and relative cell counts of CGRP- and TRPV1-positive cells in ipsilateral L3, L4, L5, L6, and S1 spinal levels of the experimental groups vs. L5 spinal level of the sham-operated group were calculated in percentage.
Statistical analysis
Characteristics and outcomes were summarized using descriptive analysis; quantitative variables are presented as means and SDs and qualitative variables as frequencies and percentages. Group comparisons of pain evaluations and of the expressions of Iba1, GFAP, CGRP, and TRPV1 were analyzed using one-way ANOVA assuming normality or using the Kruskal Wallis test without assuming normality. Multiple comparisons were performed using the Scheffe method. Comparison of pain evaluation results, expressions of Iba1, GFAP, CGRP, and TRPV1 in the NP-exposed group and sham-operated group were analyzed using the two sample t-test assuming normality or using the Mann Whitney U-test without assuming normality. P values are shown for statistically significant variables. All tests were 2-sided and P values of<0.05 were significance. The analysis was conducted using IBM SPSS ver. 19.0.
RESULTS
Pain evaluation
At 10 days after surgery, mechanical allodynia of ipsilateral hind paws was significantly lower in the NP-exposed group than in the sham-operated group (P<0.001), and mechanical withdrawal thresholds were significantly lower on ipsilateral sides in the significant pain subgroup than in the no significant pain subgroup (P<0.001).
Microglia, astrocytes, CGRP, and TRPV1
Multisegmental expressions of Iba1 and CGRP (as determined by immunochemistry) were higher in dorsal horns and DRGs, respectively, in the NP-exposed group than in the sham-operated group (Fig. 2 and 3). In the NP-exposed group, Iba1 positive microglial expression was significantly greater in L5 (P=0.004) and in ipsilateral L4 (P=0.009), L6 (P=0.002), and S1 (P=0.002) dorsal horns than in the sham-operated group. Furthermore, more Iba1 positive microglia were noted in the L5 dorsal horn and in the L4, L6, and S1 dorsal horns in the significant pain subgroup than in the no significant pain subgroup, but this difference was not statistically significant. Fig. 2 shows relative area fractions of Iba1 in ipsilateral L3, L4, L5, L6, and S1 dorsal horns vs. L5 dorsal horns in the sham-operated group. Percentage Iba1 immunoreactivities in the significant pain subgroup were 301% at L3, 397% at L4, 532% at L5, 507% at L6, and 409% at S1 dorsal horns vs. L5 dorsal horns in the sham-operated group. Percentages of CGRP positive DRG cells were also significantly higher in L5 and in ipsilateral L3, L4, L6, and S1 DRGs in the NP-exposed group than in the sham-operated group (P<0.001). Moreover, in the significant pain subgroup, the numbers of CGRP-positive cells in L5 DRGs were significantly higher than in the no significant pain subgroup (P<0.001). In addition, CGRP was up-regulated in L3, L4, L6, and S1 DRGs in the significant pain subgroup vs. the no significant pain group, but this was not significant. Relative cell counts of CGRP-positive cells at S1 DRGs in the significant pain subgroup were 866% at L3, 1,343% at L4, 1,798% at L5, 747% at L6, and 586% at vs. L5 DRGs in the sham-operated group (Fig. 3). However, the expressions of GFAP and TRPV1 were not different in the NP-exposed and sham-operated groups. Immunohistochemical examinations of GFAP in spinal cords and of TRPV1 in DGR at day 10 after surgery also revealed no significant difference between the significant pain and no significant pain subgroups (Fig. 4 and 5).
DISCUSSION
This study, which was conducted using a rat model of lumbar disc herniation, shows multisegmental upregulation of Iba1 positive microglia in dorsal horns and of CGRP expression in DRGs in the NP-exposed group as compared with the sham-operated group. In particular, in the significant pain subgroup, CGRP expression at L5 DRG was significantly higher than in the no significant pain subgroup. Moreover, it was also found that Iba1 positive microglia numbers were higher in ipsilateral L5 dorsal horns and ipsilateral L4, L6, and S1 dorsal horns, and that CGRP expression was greater in ipsilateral L5 DRGs and in ipsilateral L3, L4, L6, and S1 DRGs in the significant pain subgroup
than in the no significant pain subgroup; however, these difference were not significant.
Lumbar disc herniation is one of the most important health problems and negatively affects quality of life. Its ramifications can lead to days lost at work, activity limitations, social isolation, sleeping disorders, and intense pain (13). However, occasionally clinical motor (weakness and atrophy), sensory (pain or paresthesias), and reflex (diminution or absence of tendon reflexes) symptoms are not confined to the affected spinal cord segment as determined by MRI. Therefore, it is sometimes difficult to determine the precise location of an affected spinal cord segment, which causes difficulties creating treatment plans.
Radicular pain caused by disc herniation is mediated by biochemical and mechanical factors (13), and it has been proposed that cytokines and chemokines play major roles in the pathomechanisms of radicular pain (514). Kobayashi et al. (9) observed morphologic changes in the dorsal horn of the lumbar cord induced by nerve root compression by light and electron microscopy, and demonstrated that compressive nerve root damage is not restricted to sites of compression, but extends to synapses in the dorsal horn. Shubayev et al. (15) described the dynamics of TNFα axonal transport in the peripheral neural axis. Biotinylated TNFα was transported intra-axonally toward the periphery of normal and injured nerves, and reached the dorsal horns of spinal cords in injured rats. In the present study, we aimed to determine whether there is a neural pathway between adjacent segments of the lumbar spinal cord and DRG in a rat model of radiculopathic pain, and to provide an explanation for mismatches between symptoms and imaging results.
Microglia are the resident macrophages of the CNS and contribute to the development of chronic neuropathic pain by releasing a variety of mediators, including proinflammatory cytokines and chemokines that influence pain signaling (1617). Others have suggested nucleus pulposus application may induce glial activity in the spinal cord, and that these activated glia may play a crucial role in pain transmission in the spinal dorsal horn (711). In the present study, numbers of Iba1 positive microglia were higher in the ipsilateral L5 dorsal horns of the NP-exposed group than in the sham-operated group at 10 days after nucleus pulposus implantation. Interestingly, in addition to the L5 dorsal horn (the level of implantation), Iba1 positive microglia numbers were obviously elevated in L4, L6, and S1 dorsal horns following nucleus pulposus implantation to L5 nerve roots. Although, the significant pain and no significant pain subgroups were not significantly different in terms of Iba1 expression, numbers of Iba1 positive microglia appeared to be higher in L5 dorsal horns and in L4, L6, and S1 dorsal horns in the significant pain subgroup, and similar results were obtained for CGRP. In the NP-exposed group, CGRP expression was significantly higher in L5 DRGs and in L3, L4, L6, and S1 DRGs than in the sham-operated group. In addition, CGRP was significantly higher in L5 DRGs and non-significantly higher at the other levels in the NP-exposed group. Recently, it was reported that the effects of nucleus pulposus on nerve roots are probably due to the actions of cytokines, such as, TNFα and COX-2 (218). TNFα induces the productions of the inflammatory neuropeptides, SP, and CGRP, which are released in the spinal dorsal horn from peripheral terminals (1920). CGRP is a marker of sensory neurons primarily involved in pain perception. These results encourage us to suggest CGRP was upregulated in DRGs by cytokines and growth factors in our disc herniation model and that it is then transported into DRGs at other lumbar levels.
Therefore, we suggest that multisegmental molecular changes exist between adjacent segments of the lumbar spinal cord and DRGs in the presence of radicular pain. Fukuoka et al. (21) reported the expressions and levels of brain-derived neurotrophic factor (BDNF) and nerve growth factor (NGF) were increased in uninjured L4 DRGs after L5 spinal nerve ligation (a model of neuropathic pain), and suggested that BDNF, which was found to be increased in uninjured L4 DRG neurons, acts as a sensory neuromodulator in the dorsal horn and contributes to thermal hyperalgesia, and furthermore, that locally synthesized NGF contributes to thermal hyperalgesia. More recently, Li et al. (22) reported that after right C7 nerve root rhizotomy, c-Fos and c-Jun were expressed in the spinal gray matter of ipsilateral C7 and of ipsilateral C5 and C6, and suggested that afferent sensory fibers of the C7 nerve root project to the dorsal horn of the ipsilateral C7 spinal cord and to adjacent segments. Our results corroborate the findings of these previous studies, and show Iba1 and CGRP are up-regulated in L5 (the implantation level) and in ipsilateral L3, L4, L6, and S1. We suggested molecular changes of adjacent lumbar dorsal horns and DRGs occur because synaptic homeostasis has been disrupted. It has also been reported that glial changes following peripheral nerve injury are associated with increased sprouting of primary afferent nociceptive fibers (C and A-δ fibers) entering the spinal cord (23), morphological changes in nerve myelination and in the architecture of dorsal root ganglia (24), and the down-regulation of glial amino acid transporters (25). Furthermore, it has been suggested these morphologic and molecular changes are strictly correlated with neuro-glial plasticity changes and peripheral sensitization, and produce adaptive plasticity that facilitates neuropathic pain transmission (2627). Moreover, superficial laminae of dorsal horns of the spinal cord represent a nodal point for the modulation and integration of peripheral sensory stimuli through complex networks involving glutamate receptors and local inhibitory GABAergic interneurons (28). Cirillo et al. (29) found that the onset of reactive gliosis following peripheral nerve injury (as indicated by increases in Iba1 and GFAP) was paralleled by remarkable changes in glial and neuronal neurotransmitter transporters, as evidenced by reductions in glial glutamate and glycine transporters. The authors suggested in the presence of peripheral nerve injury reduced glutamate and glycine uptake might be responsible for the failure of the two main neuroprotective functions of neuroglia in tripartite synapses, that is, clearing excess glutamate from synaptic clefts and providing neurons with glutathione (the major antioxidant system). The findings of the present study also indicate peripheral nerve injury is able to perturb spinal synaptic circuitry by causing profound morphofunctional changes that affect neuro-glial interactions and the glutamatergic/GABAergic system (2529). Furthermore, our findings suggest perturbation of synaptic homeostasis is not always localized to spinal injury levels and can extend to adjacent spinal segments.
In the present study, we found no significant difference between GFAP immunointensities in the dorsal horns of the NP-exposed and sham-operated groups, and GFAP-positive astrocyte numbers were not significantly different in the significant pain and no significant pain subgroups. Studies using various animal models of neuropathic pain have reported astrocyte activation and GFAP expression (3031). Recently, it was suggested that injury-induced GFAP upregulation helps to maintain neuropathic pain (31). However, in a previous study, Li et al. (32) explored GFAP expression in bilateral L5 DRGs and spinal cords using immunohistochemistry after nucleus pulposus application to left L5 DRGs. They observed GFAP-immunoreactive astrocytes in bilateral spinal cord dorsal horns and found no significant difference between GFAP expressions in their nucleus pulposus and sham groups or in ipsilateral and contralateral DRGs. It also reported that spinal astrocytic proliferation did not occur after nerve root injury (33) and sciatic nerve transection (34), further supporting variation in spinal astrocytic responses in neuropathic pain model. Moreover, in a previous study, we found reactive astrocytes with thickened processes in the dorsal horn after nucleus pulposus implantation were not affected by pulsed radiofrequency administration, although mechanical allodynia was significantly attenuated (35). TRPV1 receptors have been shown to be molecular integrators of nociceptive stimuli at peripheral nerve endings, but their roles in the modulation of synaptic transmission at the spinal cord level remain unresolved (36). TRPV1 has been localized to small-diameter, unmyelinated C-fibers and medium-diameter, thinly myelinated A-δ fibers in DRGs (3738). Furthermore, spinal TRPV1 receptors have been shown to play important roles in the modulation of nociceptive transmission, especially under pathological conditions (3940). In the present study, TRPV1 expression was not significantly higher in L3, L4, L5, L6, and S1 DRGs of the NP-exposed group than in those of the sham-operated group, and no significant difference was observed between TRPV1 expressions in ipsilateral DRGs in the significant pain and no significant pain subgroups. Thus, we suggest astrocytes and TRPV1 do not play a major role in the molecular changes underlying pain-related behavior in our rat model of disc herniation.
In conclusion, our results corroborate previous reports by showing molecular changes in adjacent spinal cord segments in a model of radicular pain. To the best our knowledge, this is the first report to demonstrate multisegmental changes in terms of microglial activity in dorsal horns and CGRP expression in DRGs in lumbar spinal segments. However, we examined molecular changes only ipsilateral to a lumbar lesion and at 10 days after surgery. To determine multisegmental neural changes are responsible for these findings, studies are required to characterize molecular changes in more cranial spinal cord segments and the time courses of these changes. Our results suggest increased microglial activation and the up-regulation of CGRP expression at nucleus pulposus implantation sites and in adjacent spinal dorsal horns and DRGs. Accordingly, we suggest more detailed studies be undertaken on the mechanism responsible for these molecular changes in adjacent spinal cord segments.

 XML Download
XML Download