

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Primary carcinoma of the lung is one of the leading causes of cancer-related deaths worldwide. Lung cancer is a heterogeneous disease that can be classified into small cell lung cancer and non-small cell lung cancer (NSCLC); NSCLC accounts for about 80% of cases of lung cancer worldwide. Although the 5-yr survival rate (15%) has improved due to advances in combined modality treatment, the overall prognosis of NSCLC is still very poor (1). Thus, new treatment modalities based on the pathogenesis of NSCLC are needed.
Aberrant regulation of developmental signaling pathways is a major concern in human cancers. Mutations in the Ki-Ras gene, loss of function in tumor suppressor genes (Rb, p53), and overexpression of epidermal growth factor receptor family members are observed in NSCLC (2, 3, 4, 5, 6). These findings indicate that genetic alterations and disruptions contribute to the development of NSCLC.
There is a great deal of evidence that dysregulation of the Wnt pathway is involved in human pathologies, including cancers. The Wnt family of 19 secreted glycoproteins is highly conserved in mammals and is involved in cell fate specification, proliferation, polarity, and migration (7). APC, β-catenin, and Wnt pathway components are frequently mutated in human colon cancers (8). The Wnt pathway is also involved in other types of cancer, including hepatocellular carcinoma, breast cancer, and uterine carcinoma (9, 10, 11). Several studies indicated that Wnt7a acts as a tumor suppressor in NSCLC. Aberrant Wnt signaling has also been reported in lung cancer, and loss of Wnt7a expression occurs in some NSCLC cell lines (12, 13). Re-expression of Wnt7a shows anti-tumorigenic effects in human lung cancer cell lines through E-cadherin restoration (14). E-cadherin keeps intercellular integrity and prevents metastasis. Therefore, E-cadherin has been known as a prognostic factor in cancer. These studies indicated that Wnt7a acts as a tumor suppressor in NSCLC. However, the actual role of Wnt7a in the regulation of tumor formation in primary human NSCLC remains to be clarified.
Other than genetic alterations, epigenetic alteration is clearly acknowledged as an equally important mechanism for carcinogenesis (15). Epigenetic mechanisms consist of DNA methylation, nucleosome positioning, histone modifications, and small, noncoding RNAs (miRNA, siRNA) regulation. From these mechanisms, especially DNA methylation of the 5'-CpG island (promoter) is a major cause of tumor suppressor gene inactivation in cancers, including NSCLC (15, 16). And, the silencing of Wnt7a expression induced by promoter methylation has been identified in pancreatic cancer (17, 18).
In the present study, we performed methylation-specific PCR to examine the methylation status of Wnt7a in primary NSCLC and analyzed the correlations between aberrant Wnt7a methylation and clinicopathological factors or E-cadherin expression. Furthermore, we tried to investigate the possible effect of Wnt7a methylation on the patients' survival.
MATERIALS AND METHODS
Patients and tissue samples
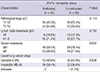
A total of 121 patients diagnosed with primary NSCLC by pathologic confirmation at Hanyang University Hospital from January 1998 to December 2000 were included in this study. Patients diagnosed only by cytology were excluded because methylation specific PCR (MSP) and immunohistochemical staining using cytology samples could not be performed. None of the patients received radiotherapy or chemotherapy before diagnosis. All diagnoses were based on pathological evidence from tissues obtained by surgery or bronchoscopic biopsy. All tissue samples were fixed in formalin and embedded in paraffin blocks. Histological classification was conducted according to World Health Organization International Histological Classification of Lung Tumor criteria (19). The tumor stage was determined according to the TNM staging guidelines suggested by the International Union Against Cancer. The demographic and clinical characteristics of the patients are summarized in Table 1.
Immunohistochemical staining
An E-cadherin immunohistochemical analysis was conducted on formalin-fixed paraffin-embedded tissue sections. Tissue sections were dehydrated in xylene and rehydrated in a graded ethanol series. Endogenous peroxidase activity was blocked by immersing the sections in hydrogen peroxide solution. The cut sections were boiled for 60 min in citrate buffer (pH 6) for heat-induced epitope retrieval. The primary antibody to E-cadherin (Cell Signaling Technology, Danvers, MA, USA) was used at a 1:100 dilution for 1 hr at room temperature. Slides were incubated for 30 min at room temperature with normal rabbit IgG at the same concentration as the primary antibody. This was followed by applying streptavidin horseradish peroxidase enzyme complex and diaminobenzidine chromogen. The slides were then counterstained in hematoxylin and coverslipped.
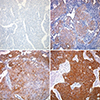
The tissue sections were scored semiquantitatively for membrane and cytoplasmic staining by light microscopy. Labeling scores were determined by multiplying the percentage of positive tumor cells per slide (0% to 100%) by the dominant staining intensity (0=negative, 1=trace, 2=weak, 3=intermediate, 4=strong). Resulting scores ranged from 0-400. Specimens with overall scores of 5-100, 101-200, 201-300, and 301-400 were classified as trace, low-level, intermediate-level, and high-level expression, respectively. Specimens exhibiting a complete absence of staining or faint staining in less than 5% of the cells were classified as negative (Fig. 1). All samples were evaluated and scored by specialized pathologist randomly without knowledge of the patient's clinical information.
Wnt7a gene promoter PCR methylation
Formalin-fixed paraffin-embedded samples were deparaffinized with standard xylene and hydrated through a graded alcohol series into water. Then, genomic DNA was extracted using a QIAamp® DNA FFPE Tissue kit (Qiagen, Valencia, CA, USA), according to the manufacturer's instructions, and stored at -80℃ until use.
Tumor DNA (100 ng) was mixed with 2 µg of salmon sperm DNA and NaOH to a final concentration of 0.3 M NaOH. Denaturation was conducted for 15 min at 37℃. Bisulfate conversion was performed with 5 M bisulfate (Zymo Research, Orange, CA, USA) solution and 100 mM hydroquinone (Zymo Research) (pH 5) at 50℃ for 6 hr. DNA modification was completed by NaOH treatment for 10 min at room temperature, followed by ethanol precipitation. The DNA was resuspended in 30 µL of sterile deionized water and used immediately or stored at -80℃.
The Wnt7a gene promoter methylation status was determined by methylation-specific PCR using bisulfite genomic DNA as the template. Two sets of primers described previously (19) were used to discriminate between the methylated and unmethylated alleles. The sense and antisense primers for the methylated Wnt7a were 5'-GTA GTT CGG CGT CGT TTT AC-3' and 5'-CGA AAC CGT CTA TCG ATA CG-3', respectively, and the primers for unmethylated Wnt7a were 5'-TAG TTT GGT GTT GTT TTA TGT TG-3' and 5'-CCC CAA AAC CAT CTA TCA ATA C-3', respectively. The PCR mixture contained 2.5 µL of 10× reaction buffer (100 mM Tris-HCl, 500 mM KCl, 15 mM MgCl2), 2 µL of deoxynucleotide triphosphates (final concentration 200 µM each), 15 pM of each primer (Cosmo, Seoul, Korea), 10 µL of modified DNA, 0.75 U of Taq™ polymerase (Intron Biotechnology Inc., Seongnam, Korea), and was adjusted with water to a final volume of 25 µL. The PCR conditions were as follows: for unmethylated DNA, 35 cycles of 95℃ for 1 min, 60℃ for 1 min, and 72℃ for 1 min; for methylated DNA, 35 cycles of 95℃ for 1 min, 58℃ for 1 min, and 72℃ for 1 min. PCR products were resolved on 2% agarose gels and visualized by ethidium bromide staining under UV illumination. In vitro methylated genomic DNA (Chemicon International, Temecula, CA, USA) was used as the positive control. A water blank was used as a negative control.
Statistical analysis
The chi-square test was used to analyze the possible correlations between clinical factors and Wnt7a methylation status of tumors. All analyses were performed using IBM SPSS statistics for Windows (IBM SPSS 21.0). In all analyses, P<0.05 was taken to indicate statistical significance.
RESULTS
Methylation frequency of the Wnt7a gene promoter in lung cancer tissue samples
A methylation-specific PCR (MSP) assay was performed to determine the methylation status of the Wnt7a gene promoter in 121 NSCLC tissues. Fig. 2 shows representative MSP results. Of the 121 tumor tissue samples collected from patients with primary NSCLC, 32 (26.4%) showed methylation in the Wnt7a gene promoter region. We also performed MSP of Wnt7a in non-tumor lung tissue (50 of 121 samples) obtained from surgically resected specimens of patients with lung cancer. None of the 50 (0%) corresponding non-tumor tissue samples showed methylation in the Wnt7a promoter region.
Correlations between aberrant Wnt7a methylation and clinicopathological factors
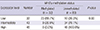
We analyzed the associations between Wnt7a gene promoter methylation status and various clinicopathological characteristics of patients with NSCLC, including gender, age, histological type, smoking habit, and TNM staging. Because the acquisition of the clinical data was performed with retrospective chart review of more than 10 yr ago, there were inevitable missing data in the smoking status, pathologic T or N stage (Table 1). As shown in Table 2, Wnt7a promoter methylation showed no significant correlation with baseline demographic factors like gender, age and smoking status. Even though local tumor progression and lymph node metastasis status also showed no significant correlation with Wnt7a promoter methylation, patients with distant metastases showed higher frequency of Wnt7a promoter methylation than the patients without metastases as shown in Table 3 (40.6% vs. 21.6%, P=0.037). Moreover, the rate of Wnt7a methylation was significant higher in patients with a surgically inoperable (IIIb and IV) tumor stage than operable stage (37.3% vs. 18.8%, P=0.036).
Correlation between Wnt7a methylation and E-cadherin expression
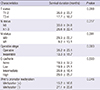
Wnt7a methylation was preferentially observed in cases with low E-cadherin expression (Table 4). The frequency of Wnt7a methylation was 69.7% in patients with trace or low E-cadherin expression. Furthermore, no Wnt7a promoter methylation was observed in patients with high E-cadherin expression level. As a result, very strong negative correlation was found between E-cadherin expression and Wnt7a promoter methylation. It means that Wnt7a methylation would cause the loss of E-cadherin expression.
Correlation between Wnt7a methylation and survival of the patients
We could confirm the survival status of 71 patients with retrospective chart review and available patients contact. We analyzed the possible correlation between Wnt7a methylation and long-term survival. Univariate analysis showed that presence of Wnt7a methylation, inoperable stage or higher T and M stage had significantly shorter survival (Table 5).
However, with Kaplan-Meyer curve with available survival status and cox regression model including sex, age, smoking status, stage, and E-cadherin expression, there was no significant correlation between Wnt7a methylation and survival (Fig. 3).
DISCUSSION
In this study, we detected Wnt7a promoter methylation status in patients with NSCLC and analyzed its association with clinicopathological factors of lung cancer. Overall, 26.4% of patients with NSCLC showed Wnt7a promoter methylation. Wnt7a promoter methylation was strongly correlated with loss of E-cadherin expression, local tumor progression and distant metastasis, higher stage and possibly survival. The results suggested that Wnt7a methylation may be a promising prognostic factor for NSCLC. In particular, the correlation between the loss of E-cadherin and Wnt7a methylation was interesting and may be clinically useful.
E-cadherin is a direct mediator of cell - cell adhesion interactions. A reduction in intercellular adhesion is the main cause of the invasive properties of many primary malignant tumors. The cytoplasmic tail of E-cadherin is linked to the cytoskeleton via catenins. Loss of E-cadherin in cancer induces an epithelial-mesenchymal transition and is associated with cell de-differentiation, invasion, and metastasis (20). Several clinical studies have shown that reduced E-cadherin and catenin levels result in poor prognosis and short survival in patients with NSCLC (21, 22, 23, 24, 25, 26, 27, 28, 29). There are various mechanisms that may underlie a loss of E-cadherin, such as downregulation of growth factor receptors, promoter methylation, or gene mutations (30, 31). Several transcriptional repressors also inhibit E-cadherin gene expression (32, 33). In the present study, patients with NSCLC and reduced E-cadherin expression had a high frequency of Wnt7a promoter methylation. Furthermore, Wnt7a methylation was not detected in patients showing normal E-cadherin expression.
There is also evidence that E-cadherin is a Wnt target gene. In Drosophila, an E-cadherin homolog is regulated by Wnt signaling (34). In mice, the E-cadherin promoter has a LEF-1 transcriptional binding site that is activated by β-catenin (35), and Wnt/β-catenin signaling upregulates cadherin-11 expression in Xenopus (36). Several previous studies indicated the loss of Wnt7a mRNA in lung cancer cell lines and primary tumors (12, 13, 14). Ohira et al. (37) reported that E-cadherin was induced by Wnt7a transfection in a cancer cell line. In this previous study, Wnt7a was also shown to increase cytoplasmic β-catenin levels. They assumed that E-cadherin was induced through the canonical Wnt pathway. Winn et al. (14) also demonstrated that Wnt7a induces E-cadherin through the Frizzled-9 receptor. However, they did not determine whether Wnt7a stimulated β-catenin/T cell factor activity. Induction of Wnt7a by E-cadherin is mediated through ERK-5-dependent activation of PPAR-γ in NSCLC cells (14). Restoration of Wnt7a expression accompanied by induction of E-cadherin protein expression stimulates epithelial differentiation and inhibition of NSCLC cell proliferation (13). Therefore, it seems likely that loss of Wnt7a caused by promoter methylation results in the loss of E-cadherin expression in human NSCLC. While several in vitro and ex vivo studies have indicated that Wnt7a acts as a tumor suppressor, there have been no human studies to identify the clinical significance of Wnt7a.
This is the first study to analyze the relationship between loss of Wnt7a and clinicopathological status in human lung cancer. Although we did not demonstrate that the loss of E-cadherin was caused by Wnt7a methylation, a strong inverse correlation was observed between the loss of Wnt7a and E-cadherin expression. Wnt7a promoter methylation was detected in 69.7% of the patients showing trace or low-grade E-cadherin expression. Moreover, Wnt7a promoter methylation was not detected in the patients showing high grade E-cadherin expression (Table 4). It is unclear whether Wnt7a promoter methylation and loss of E-cadherin are coincident or causal, as promoter methylation underlies various gene silencing mechanisms and another pathogenic cause may be involved in the reduction of E-cadherin expression accompanied by Wnt7a methylation. Our data suggest that loss of Wnt7a plays an important role in decreased E-cadherin expression.
We also analyzed the prognostic effects of Wnt7a methylation on long-term survival. Even though the patients with Wnt7a methylation seemed to show shorter survival with univariate analysis, multivariate analysis using Cox regression model showed no significant correlation between Wnt7a methylation and long term survival. This finding could implicate that the effect of Wnt7a methylation on the prognosis of NSCLC might be not so strong as the profound effect of advanced staging or presence of metastasis. In addition, in this study, we excluded all NSCLC patients diagnosed by cytology only because methylation PCR could not be performed. Therefore, it is possible that many patients with inoperable stages were counted out, and our survival data seems to be influenced by selection of subjects.
In addition to the staging system, we observed that E-cadherin expression and Wnt7a methylation showed an inverse correlation in this study regardless of causal relation. Moreover, E-cadherin is a well-known marker of NSCLC prognosis (21). Thus, it was presumed that loss of Wnt7a by promoter methylation would be a poor prognostic factor not directly but indirectly via E-cadherin expression and the advanced staging. Because of the retrospective study design and large portion of missing survival information, further studies are necessary to clarify the usefulness of Wnt7a as a prognostic marker and a novel treatment modality. Because of the weak or less significant correlation between Wnt7a methylation and the overall survival of the patients in our study, comparison studies including Wnt7a methylation and other biomarkers including various genetic or epigenetic alteration for the prediction of the clinical course in NSCLC could be helpful.
In summary, the overall frequency of Wnt7a promoter methylation was 26.4% of human NSCLC tissues in our study. The rate of Wnt7a loss by promoter methylation was significantly higher in patients with advanced tumor stages than in those with less advanced stages. We also demonstrated the correlation between the loss of E-cadherin and Wnt7a promoter methylation. These findings suggest that loss of the Wnt7a gene induced by promoter methylation may be an important prognostic factor for NSCLC.


 XML Download
XML Download