

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
The ends of chromosomes are composed of repetitive nucleotide sequences called telomeres. The main role of telomeres is to protect chromosome from degradation, end-to-end fusion, and atypical recombination by capping chromosome ends (1). Therefore, telomeres are essential for maintaining genomic integrity and stability (2). It is known that telomeres get shorter during mitosis due to incomplete replication of linear chromosomes by conventional DNA polymerases (1). When telomeres are shortened to a critical length, chromosome instability increases and cellular senescence or apoptosis occurs (3). Abnormally shortened telomeres can lead to diverse human diseases (4567). A number of studies have also reported that short telomeres are associated with development of lung cancer (8910).
Polymorphisms in telomere biology genes, such as telomerase reverse transcriptase (TERT) and telomerase RNA component (TERC), can influence telomere length. Recently, a number of candidate gene studies and genome-wide association studies (GWAS) have identified single nucleotide polymorphisms (SNPs) that are related to telomere length in Caucasians (11121314). In a previous study, we evaluated the telomere lengths of 17 SNPs related to telomere length in Caucasians and verified that 6 SNPs (TERT rs2853669, rs2736108, rs2736098, and rs7705526; TERC rs2293607; and ZNF676 rs412658) are associated with short telomere length in the Korean population (15). Here, we conducted a case-control study to investigate the association between the 6 SNPs related to short telomere length and lung cancer risk in the Korean population.
MATERIALS AND METHODS
Study population
This study involved 1,100 lung cancer patients and 1,096 healthy controls. The patients were first diagnosed with lung cancer between January 2008 and July 2012 at Kyungpook National University Hospital (KNUH), Daegu, Korea. There were no restrictions on gender, histologic subtype, or stage. The control subjects were frequency matched (1:1) to the cases based on gender and age (±5 yr) from a pool of healthy volunteers who visited the general healthy check-up center at KNUH during the same period. All subjects were of Korean ethnicity and residing in Daegu City or the surrounding regions. Blood samples were provided by the National Biobank of Korea, which is supported by the Ministry of Health, Welfare, and Family Affairs.
SNP selection and genotyping
In a previous study, we evaluated the associations between telomere length in the Korean population and 17 SNPs related to telomere length in Caucasians (15). Six of the 17 SNPs were associated with short telomere length in the Korean population and were selected for the present case-control study. Four SNPs in TERT (rs2853669, rs2736108, rs2736098, and rs7705526), one SNP in TERC (rs2293607), and one SNP in ZNF676 (rs412658) were genotyped. The rs7705526 polymorphism was genotyped by PCR-RFLP analysis, and the others were genotyped by melting-curve analysis using fluorescence-labeled hybridization probes (LightCycler 480, Roche Diagnostic, Mannheim, Germany). Samples that could not be scored by the LightCycler were re-genotyped by direct sequencing using an ABI 3730xl sequencer (Invitrogen, Garlsbad, CA, USA). The call rates for the SNPs were more than 95% (Supplementary Table 1). All genotyping analyses were blinded with respect to the case or control status to ensure quality control.
Statistical analysis
Comparisons between cases and controls were achieved by the Student t-test for continuous variables or the chi-square test for categorical variables. Hardy-Weinberg equilibrium was assessed by a goodness-of-fit chi-square test. To estimate lung cancer risk related to the SNPs, multivariate logistic regression analysis was used to calculate the adjusted odds ratio (aORs) and 95% confidence intervals (CIs) after adjusting for age, gender, and pack-years of smoking. For stratified analysis, individuals were grouped according to age (<62 yr vs. ≥62 yr), gender (male vs. female), smoking status (never vs. ever), histologic subtype (squamous cell carcinoma vs. adenocarcinoma vs. small cell carcinoma), and stage (I or II vs. III or IV). To compare aORs in each subgroup, homogeneity was tested using the Wald method. Statistical analyses were performed using the SPSS 21.0 software (SPSS Inc., Chicago, IL, USA).
RESULTS
The demographics of the cases and controls are shown in Table 1. Significant differences in smoking status and pack-years of smoking were observed between cases and controls (P<0.001). Therefore, these differences were adjusted in the multivariate analyses. Basic information for the 6 SNPs, including gene locations and minor allele frequencies are shown in Supplementary Table 1. The genotype frequencies of the 6 SNPs were in Hardy-Weinberg equilibrium.
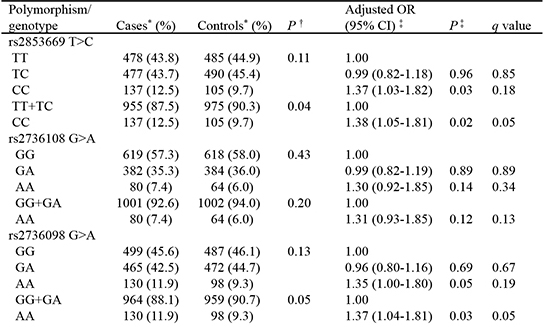
The genotype distributions of the 6 SNPs in the cases and controls are shown in Table 2. Among the 6 SNPs, the genotype frequencies of TERT rs2853669 and rs2736098 differed significantly between cases and controls. After adjusting for age, gender, and pack-years of smoking, individuals carrying the rs2853669 CC genotype had a significantly increased risk of lung cancer compared to those with the rs2853669 TT or TC genotypes (aOR=1.38, 95% CI=1.05-1.81, P=0.02). The rs2736098 G>A genotype was also significantly associated with lung cancer risk under a recessive model (aOR=1.37, 95% CI=1.04-1.81, P=0.03).
The TERT rs2853669 and rs2736098 polymorphisms were in strong linkage disequilibrium (LD) (|D'|=0.95, Supplementary Fig. 1). The rs2853669 polymorphism is known to be a potentially functional polymorphism (16). Therefore, we examined the association between TERT rs2853669 and lung cancer risk after stratifying the subjects by age, gender, smoking status, histologic subtype, and stage (Table 3). When stratified according to age, the effect of rs2853669 on lung cancer risk was significant in younger individuals (aOR=1.73, 95% CI=1.18-2.54, P=0.005) but not in older individuals (aOR=1.08, 95% CI=0.74-1.59, P=0.68; P value for homogeneity test [PH]=0.09). When stratified according to histologic subtype, significant association between rs2853669 and lung cancer risk was observed in adenocarcinoma (AC) (aOR=1.50, 95% CI=1.07-2.07, P=0.02) but not in squamous cell carcinoma (SCC) or small cell lung cancer (SCLC) (aOR=1.16, 95% CI=0.78-1.73, P=0.46; aOR=1.18, 95% CI=0.66-2.11, P=0.57, respectively).
We conducted interaction analysis between rs2853669 and smoking status. When non-smokers with rs2853669 TT or TC genotype was considered as reference group, smokers with rs2853669 CC genotype had a significantly increased risk of lung cancer (OR=3.71, 95% CI=2.35-5.86, P=1.9×10-8, Supplementary Table 2).
DISCUSSION
In this study, we investigated the association between 6 SNPs related to short telomere length in the Korean population and lung cancer risk via a case-control study. Among the 6 SNPs, two SNPs (rs2853669 and rs2736098) in TERT were significantly associated with increased lung cancer risk in the Korean population.
TERT is one of the major components of telomerase (1718). Telomerase is a reverse transcriptase that can add nucleotide sequence to telomere ends, thus protecting telomere length from progressive shortening (19). Polymorphisms in TERT can influence telomere length (162021) and can also affect cancer susceptibility (202223). In the present study, the TERT rs2736098 AA genotype had an increased risk of lung cancer compared with the TERT rs2736098 GG or GA genotypes. The rs2736098 polymorphism is located in exon 2 of TERT and is a synonymous polymorphism (A305A). In the present study, rs2736098 was strongly linked to rs2853669 (|D'|=0.95), which is located in the promoter region of TERT. TERT rs2853669 is known to be a potentially functional polymorphism (16). Therefore, the effect of rs2736098 on lung cancer risk may be due to LD with rs2853669. In the present study, individuals that had the TERT rs2853669 CC genotype had a significantly increased risk of lung cancer compared with those that had the rs2853669 TT or TC genotypes. In the previous study, relative telomere length of individual with rs2853669 CC genotype was significantly shorter than those with rs2853669 TT or TC genotype (1.97±0.6 vs. 2.37±0.6, P=0.03) (15). Increased risk of cancer in individual with short telomere is biologically plausible, especially in light of the function of telomere. Main function of telomere is to keep genomic integrity and stability from deterioration or atypical fusion of chromosome (2). Therefore, the shortened telomere can lead to chromosomal instability and may thereby increase cancer risk. Recently, it was reported that the TERT rs2853669 C allele was associated with an increased risk of lung cancer in a Chinese population (23). That the results from these two independent studies are consistent is strong evidence that TERT rs2853669 is associated with lung cancer risk.
When stratified according to histologic subtype, TERT rs2853669 was associated with lung cancer risk in AC but not in SCC or SCLC. Zhong et al. (23) also reported that increased risk of lung cancer due to a variant of rs2853669 was more prominent in AC. Although AC and SCC are categorized as NSCLC, AC has a different molecular pattern, carcinogenesis, and treatment response from SCC (242526). Differential expression of TERT between lung AC and SCC or SCLC was also reported in a previous study (27). Therefore, TERT rs2853669 may influence lung cancer susceptibility differently according to histologic subtype. Additional studies are required to confirm this finding.
Interestingly, the effect of TERT rs2853669 on lung cancer risk was different by age. TERT rs2853669 was associated with a significantly increased risk of lung cancer in younger individuals but not in older individuals; this difference showed a borderline significance (PH=0.09). This suggests that short telomere length due to TERT rs2853669 could play a predominant role in the development of lung cancer at earlier ages.
Among the 6 SNPs related to short telomere length in the Korean population, only two SNPs (TERT rs2736098 and rs2853669) were associated with lung cancer risk. The TERT rs2736108 AA genotype tended to associate with increased lung cancer risk, but the association was not statistically significant. TERT rs7705526, TERC rs2293607, and ZNF676 rs412658 were not associated with lung cancer risk. Since carcinogenesis is a multistep process (28), multiple genetic and epigenetic changes are required to transform a normal cell into a cancer cell (29). A mouse model with shortened telomeres and mutant p53 showed increased hepatocellular carcinoma progression, while a model with shortened telomeres and functional p53 did not (30). Therefore, a single nucleotide change that influences telomere length may not have sufficient power to affect lung cancer risk. Additionally, a polymorphism that causes short telomere length may influence lung cancer risk separately from telomere length, and so the increased risk of lung cancer due to short telomere length can be weakened by another biologic effect of the polymorphism.
In summary, we previously investigated the associations between lung cancer risk and 17 SNPs related to telomere length in Caucasians. We identified 6 SNPs associated with short telomere length in the Korean population. Among these, TERT rs2853669 and rs2736098 were associated with increased lung cancer risk in the present study. This suggests that a common functional promoter polymorphism, TERT rs2853669, may contribute to both telomere length and lung cancer risk in the Korean population.




 XML Download
XML Download