

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Ubiquitin, which is a protein modifier consisting of 76 amino acids, plays a key role in protein degradation or in the alteration of protein function. The ubiquitin binding to a lysine residue is achieved through the actions of three classes of enzymes, namely ubiquitin-activating enzyme (E1), ubiquitin-conjugating enzyme (E2), and ubiquitin ligase (E3) (1). When ubiquitin conjugates with proteins as a monomer (mono-ubiquitination), it determines the subcellular localization of the proteins or modulates the binding affinities of the proteins to their partner proteins and target DNAs (2). Ubiquitins also conjugate with proteins as a polymer form, which is called "poly-ubiquitination", and the poly-ubiquitinated proteins undergo degradation through proteasomes (3). As a huge amount of proteins is regulated through ubiquitination, cells should cope with the rapid consumption of free ubiquitin. Indeed, the cellular pool of free ubiquitin is maintained through two pathways, de novo synthesis and recycling. The ubiquitin pool is replenished by deubiquitinases, which recycle ubiquitin monomers by cleaving polymerized ubiquitin chains clung to proteins during proteasomal degradation. To date, three deubiquitinases (PSMD14, USP14 and UCHL5) in association with the proteasome have been known to be involved in the ubiquitin recycling (45).
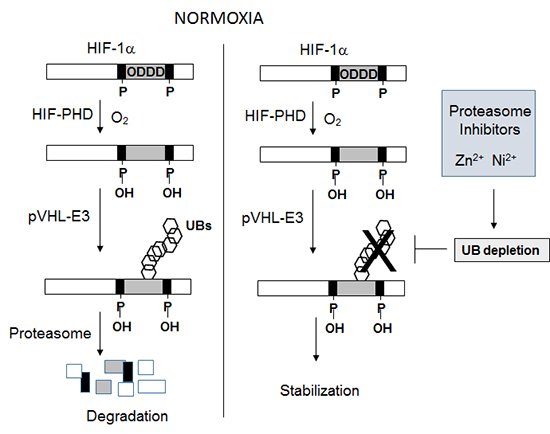
Hypoxia-inducible factor 1alpha (HIF-1α), which is detected approximately at 120 kDa on Western blotting, is a representative protein whose fate is mainly determined through ubiquitination (6). HIF-1α acts as a master regulator to activate the hypoxia-induced genes essential for cellular response to hypoxia (7). Under normoxic conditions, HIF-1α is hydroxylated at two proline residues in the oxygen-dependent degradation domain (ODDD) by HIF prolyl-hydroxylases and this modification induces the poly-ubiquitination of HIF-1α through the pVHL-E3 ligase complex, thereby leading to the proteasomal degradation of HIF-1α (8). Under hypoxic conditions, however, HIF-1α is stabilized because the prolyl-hydroxylation is limited.
According to the above process for HIF-1α regulation, it seems to be unquestionable that HIF-1α is upregulated even under normoxic conditions by the inhibition of proteasome, which has been evidently demonstrated in many literatures (9). In this study, we had a question about why HIF-1α is stabilized by proteasome inhibition. Considering that only poly-ubiquitinated HIF-1α can be targeted by the proteasomes, it is so reasonable that poly-ubiquitinated HIF-1α is accumulated by proteasome inhibition. Moreover, if the proteasome inhibition affects neither HIF-1α prolyl-hydroxylation nor HIF-1α ubiquitination process, the major portion of proteasome inhibition-stabilized HIF-1α is expected to be poly-ubiquitinated and then to be detected at a higher level than 120 kDa. However, many researchers empirically have known that there are no substantial differences in molecular weight on immunoblotting between the proteasome inhibition-stabilized HIF-1α and the hypoxiainduced, non-ubiquitinated HIF-1α (10). To unravel this mystery, we tested the possibility that proteasome inhibition blocks the process for poly-ubiquitination of HIF-1α. Our results suggest that under proteasome inhibition, the ubiquitin pool is depleted due to the impairment of ubiquitin recycling and by doing so HIF-1α is not poly-ubiquitinated under normoxia.
MATERIALS AND METHODS
Reagents and antibodies
Culture media were purchased from Invitrogen (Carlsbad, CA, USA), and fetal bovine serum from Sigma-Aldrich (St. Louis, MO, USA). Anti-HIF-1α antibody was raised in rabbits against the ODDD peptide of human HIF-1α (11). Antibodies against ubiquitin (sc-8071), α-tubulin (sc-9104), and HA tag (sc-805) were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA), anti-GFP (2555) from Cell Signaling (Danvers, MA, USA), and anti-FLAG tag (F7425) from Sigma-Aldrich. MG132 was purchased from Enzo Life Science (Farmingdale, NY, USA), and lactacystin and bortezomib from Santa Cruz Biotechnology. Zinc chloride and nickel chloride were purchased from Sigma-Aldrich, and cobalt chloride from Junsei Chemical Co. (Tokyo, Japan).
Cell lines and culture
HEK293T (human embryonic kidney), HCT116 (human colorectal carcinoma), and SiHa (human squamouse cell carcinoma) cell-lines were obtained from American Type Culture Collection (Manassas, VA, USA). Cell lines were cultured in Dulbecco's modified Eagle's medium (DMEM) containing 10% fetal bovine serum at 20% O2/5% CO2 for normoxic incubation or at 1% O2/5% CO2 for hypoxic incubation.
Preparation of plasmids and transfection
The cDNAs of ubiquitin, HIF-1α, HIF-1α ODDD (aa. 394-704), and IκBα were cloned by RT-PCR and PCR using the pfu DNA polymerase, and inserted into hemagglutinin (HA)-tagged, GFP-tagged, or FLAG-tagged pcDNA plasmid. Cells at ~40% density were transiently transfected with the plasmids using calcium phosphate or Lipofectamine 2000 reagent (Life Technologies; Grand Island, NY, USA). The transfected cells were allowed to stabilize for 48 hr before experiments.
Immunoblotting and immunoprecipitation
For immunoblotting, total proteins in cell lysates were electrophoresed on SDS-polyacrylamide (8% or 15%) gels, and transferred to a PVDF membrane (Millipore; Bedford, MA, USA). Membranes were blocked with a Tris/saline solution containing 5% skim milk and 0.1% Tween-20 for 1 hr and incubated overnight at 4℃ with a primary antibody (1:1,000) in the blocking solution. The membranes were incubated with a horseradish peroxidase-conjugated secondary antibodies (1:5,000) for 1 hr at room temperature. Proteins were visualized using an ECL Prime (GE healthcare, Pittsburgh, PA, USA). To analyze protein interaction, total protein extracts were prepared using a cell lysis buffer containing 1% NP-40 and protease inhibitor cocktails (Sigma-Aldrich). Total proteins were incubated with anti-FLAG affinity beads (Sigma-Aldrich) for 4 hr at 4℃. Bound proteins were eluted in a denaturing SDS sample buffer, and subjected to immunoblotting.
Quantitative RT-PCR (RT-qPCR)
RNAs were isolated from cells using TRIzol® reagent (Life technologies). cDNAs were synthesized from 1 µg of RNAs using Tetro cDNA synthesis Kit provided by Bioline (Taunton, MA, USA). The cDNAs were amplified using the SensiFAST™ SYBR® No-ROX Kit (Bioline). Real-time polymerase chain reaction were performed in 96 well optical plates, data were analyzed using CFX manager software (Bio-Rad laboratories; Hercules, CA, USA) and the values were normalized to the GAPDH levels.
RESULTS
Comparison of molecular weights among HIF-1α proteins induced by proteasome inhibitors and hypoxia
As was expected, HIF-1α protein was stabilized either under hypoxia or by a proteasome inhibitor MG132 under normoxia. Interestingly, HIF-1α proteins stabilized under both conditions were detected at the same level (120 kDa) on a denaturing SDS/PAGE in HEk293T, HCT116, and SiHa cell lines (Fig. 1A). We checked the time course of HIF-1α stabilization by MG132, and found that HIF-1α is overtly induced after 2 hr-incubation in three cell lines. To understand why the stabilized HIF-1α is not poly-ubiquitinated, we first checked the cellular level of free ubiquitin in the presence of MG132. The ubiquitin level was strikingly decreased in three cell lines after 2 hr-incubation with MG132 (Fig. 2B). Likewise, HIF-1α was induced but the ubiquitin level was reduced by other proteasome inhibitors lactacystin and bortezomib (Fig. 1C). These results suggest that the ubiquitin pool is significantly reduced within 2 hr after proteasome inhibition and then non-ubiquitinated HIF-1α proteins are accumulated at a later period.
Effect of free ubiquitin restoration on HIF-1α accumulation under proteasome inhibition
If HIF-1α is not poly-ubiquitinated due to the depletion of the ubiquitin pool under proteasome inhibition, the HIF-1α ubiquitination might be resumed by restoration of free ubiquitin. We first checked whether ectopically expressed HA-ubiquitin participates in the ubiquitination of endogenous proteins. The levels of ubiquitinated proteins were increased by the expression of HA-ubiquitin (Fig. 2A), which verifies that expressed HA-ubiquitin functions to conjugate with endogenous proteins. We next transfected three cell lines with HA-ubiquitin, and checked the HIF-1α levels in the presence of MG132. The cellular levels of endogenous HIF-1α were substantially reduced in three cell lines expressing HA-ubiquitin (Fig. 2B). Even ectopic GFP-HIF-1α was stabilized by MG132, which was attenuated by HA-ubiquitin expression (Fig. 2C). To make sure that the poly-ubiquitination of HIF-1α is resumed by restoration of free ubiquitin, we expressed FLAG-HIF-1α ODDD in HEK293T cells and analyzed the cellular level of poly-ubiquitinated ODDD. The poly-ubiquitination of HIF-1α ODDD was profoundly stimulated by the expression of HA-ubiquitin (Fig. 2D). Taken together, these results strongly support our notion that the HIF-1α stabilization under proteasome inhibition is attributed to the depletion of the ubiquitin pool. As the depletion of the ubiquitin pool under proteasome inhibition may affect the ubiquitination of proteins other than HIF-1α, we tested the possibility that the poly-ubiquitination of IκB-α is resumed by restoration of free ubiquitin. Subsequently, IκB-α was also stabilized by MG132, which was reversed by HA-ubiquitin expression. Moreover, IκB-α was poly-ubiquitinated by HA-ubiquitin even under proteasome inhibition (Fig. 2E). This suggests that the accumulation of non-ubiquitinated proteins under proteasome inhibition is not restricted to HIF-1α.
Depletion of the ubiquitin pool by metal ions and HIF-1α stabilization
It is well known that the normoxic stabilization of HIF-1α can be induced under various stress conditions. Indeed, transition metal ions, such as Co2+, Ni2+, and Zn2+, have been widely used as hypoxia-mimic agents capable of inducing HIF-1α (111213). In a view point of mechanism, these agents has been considered to inhibit the prolyl-hydroxylation of HIF-1α because the enzymatic reaction essentially requires Fe2+ as a cofactor (8). The transition metal ions may inhibit the hydroxylation process by competing with Fe2+. However, the precise actions of these agents have not been elucidated so far. Therefore, we tested the possibility that the hypoxia-mimic agents stabilize HIF-1α through the depletion of the ubiquitin pool. Surprisingly, the cellular level of free ubiquitin was reduced by Ni2+ and Zn2+, but not by hypoxia and Co2+ (Fig. 3A). Moreover, HIF-1αinduction by Ni2+ and Zn2+ was significantly attenuated by restoration of free ubiquitin, whereas HIF-1α induced by hypoxia and Co2+ was not (Fig. 3B). The depletion of the ubiquitin pool may contribute to HIF-1α induction in part by Ni2+ and Zn2+. To rule out the possibility that the ubiquitin level is reduced at the translational level, the mRNA levels of three ubiquitin genes (UBA, UBB, and UBC) were analyzed using RT-qPCR. All of three gene transcripts were found to be significantly induced in the presence of MG132, ZnCl2, or NiCl2 (Fig. 4A). The mRNAs of ubiquitins seem to be induced to replenish the ubiquitin pool, and thus, the ubiquitin depletion might not be attributed to impaired synthesis of ubiquitin. Possibly, the ubiquitin pool runs out under proteasome inhibition because free ubiquitins cannot be recycled. Then, do ZnCl2 and NiCl2 also block the recycling of ubiquitin? To answer this question, we examined if the metal ions increase the levels of conjugated ubiquitins. For this experime nt, FLAG-HIF-1α ODDD was used as a representative protein subjected to ubiquitin conjugation. Unexpectedly, ZnCl2 and NiCl2 both induced the accumulation of poly-ubiquitinated peptides, whereas hypoxia and CoCl2 did not (Fig. 4B). This result indicates that ZnCl2 and NiCl2 disturb the recycling of free ubiquitin, as MG132 does. In terms of mechanism underlying the metal-induced stabilization of HIF-1α, the ubiquitin depletion as well as the inhibition of prolyl-hydroxylation may contribute to the stabilization of HIF-1α.
DISCUSSION
This study was performed to answer why HIF-1α protein is normoxically accumulated as a non-ubiquitinated form in the presence of proteasome inhibitors. Consequently, HIF-1α was found not to be ubiquitinated due to the depletion of the ubiquitin pool under proteasome inhibition. We also found that transition metal ions, such as Ni2+ and Zn2+, deplete the ubiquitin pool and by doing so accumulate the non-ubiquitinated HIF-1α even under normoxia. To date, the hydroxylation-ubiquitination-degradation process has been extensively investigated to understand the oxygen-dependent regulation of HIF-1α. Based on these results, the state of the ubiquitin pool seems to be another critical factor determining the cellular level of HIF-1α. This scenario is summarized in Fig. 5.
It was previously reported that the ubiquitin pool is depleted by proteasome inhibitors. Xu et al. demonstrated that various proteasome inhibitors, such as MG132, bortezomib, lactacystin, and ALLN, rapidly reduced free ubiquitin levels in aT3-1 pituitary cells and subsequently inhibited the ubiquitin thiolesterification to E2 enzymes (14). They also found that the mono-ubiquitination of histone and the poly-ubiquitination of inositol trisphosphate receptor were blocked by the proteasome inhibitors. In addition, Melikova et al. also reported that the MG132 stabilization of epidermal growth factor receptor results from depletion of the free ubiquitin pool (15). Similarly, we found that HIF-1α is not poly-ubiquitinated and stabilized due to ubiquitin depletion under proteasome inhibition. The ubiquitin depletion might be one of major factors contributing to diverse pharmacological effects of proteasome inhibitors.
In the present study, the most striking finding is that nickel and zinc ions block the poly-ubiquitination of HIF-1α by depleting the free ubiquitin pool. Since the metal ions induced the mRNA levels of three ubiquitin isoforms (Fig. 4A), they might not inhibit the de novo synthesis of ubiquitins at the transcriptional level. Then, how do these metal ions reduce the free ubiquitin level? We can propose two possible hypotheses: 1) If the metal ions can damage preexisting proteins or induce the premature termination of protein synthesis or maturation, free ubiquitins could be consumed for ubiquitinating and degrading such useless proteins; 2) The metal ions could block the recycling of free ubiquitin via the inhibition of proteasomes. Although the former possibility could not be excluded, we can make a hasty conclusion that the later one is reasonable at least in our experimental settings. As shown in Fig. 4B, poly-ubiquitinated HIF-1α ODDD was markedly accumulated by the treatment of nickel or zinc ion. This result supports the later hypothesis that the metal ions inhibit the proteasomal degradation of poly-ubiquitinated peptides.
What is the mechanism by which the proteasome-mediated degradation of HIF-1α is interrupted by nickel and zinc ions? Two previous studies have shown that Zn2+ influx interferes with proteasome function in human cancer cells (1617). Many poly-ubiquitinated proteins were accumulated in HeLa cells treated with 250 µM ZnSO4 alone or co-treated with 1 µM ZnSO4 and 100 µM pyrithione, a zinc ionopore that helps zinc ion enter into cells (16). Such an anti-proteasomal effect of zinc was clearly demonstrated by checking fluorescence intensity of Ub(G76V)-GFP fusion protein that is easily degradable by proteasomes. Likewise, Qiu et al. reported that pyrithione inhibited proteasome function Zn2+-dependently in various human cell lines (17). In addition, Cvek et al. reported that the Zn2+-diethyldithiocarbamate complex inhibits the chymotrypsin-like activity of 20S proteasome both in test tubes and in cells (18). Given our results and these literatures, Zn2+ may deplete the ubiquitin pool through its inactivation of proteasomal enzymes, leading to the stabilization of HIF-1α. In contrast, the anti-proteasomal activity of nickel ion has not been reported so far. Frezza et al. examined whether Ni2+ or Zn2+ has an inhibitory effect on proteasome activity using a purified rabbit 20S proteasome, and found that Zn2+ effectively inactivates the proteasome but Ni2+ does not (19). Nonetheless, Ni2+ showed an inhibitory activity on the proteasomal degradation of poly-ubiquitinated proteins in our experimental settings (Fig. 4B). The effect of Ni2+ on proteasomal activity is an open question.
Transition metal ions and iron chelators are able to stabilize HIF-1α even under normoxia and thus, they have been popularly used as hypoxia-mimic agents. It is well-known that iron chelators block the oxygen-dependent hydroxylation of HIF-1α by removing Fe2+ from HIF-prolyl-hydroxylases. Given the essential role of Fe2+ in the hydroxylation process, divalent metal ions have been considered to inhibit the hydroxylation of HIF-1α because they can act competitively against divalent iron ion. Actually, this concept has be accepted without serious conflicts because the competition between transition metal ions often occurs in biological processes. We also think that such an idea is scientifically reasonable. However, our argument in this study is that depletion of free ubiquitin could be another factor contributing to divalent metal-induced expression of HIF-1α. As HIF-1α is one of the most compelling targets for treating hypoxia-associated diseases including infarctions, angiogenesis and neoplasms, it is very important to know the mechanisms underlying HIF-1α regulation more accurately. This work may provide a better understanding of HIF-1α regulation.



 XML Download
XML Download