

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Wound healing is initiated and progressed through a complex integrated sequence of cellular, physiologic, and biochemical events, such as inflammation, granulation, re-epithelialization and remodeling process. The abnormal process causes the non-healing chronic wounds or the abnormal scar and keloid (1, 2). Many biological factors, such as growth factors and cytokines, are necessary for the multiplex steps of wound healing contributed to their roles in promoting migration of inflammatory cells, fibroblasts, and endothelial precursor cells into the wound site (3, 4). The migration of keratinocytes is essential for wound re-epithelialization and re-establishment of skin remodeling (5). Activated keratinocytes could secret various multifunctional cytokines that affect to the activation of inflammatory cells during wound healing process (6, 7, 8). One of them, interleukin-6 (IL-6) is a multifunctional cytokine that regulate the immune response and wound healing in a timely manner. It was reported that IL-6 signaling pathway was involved in activation of corneal wound healing and pathogenesis of keloid formation, respectively (7, 9) In IL-6 deficient mice, IL-6 showed crucial roles in wound healing by regulating leukocyte infiltration, angiogenesis, and collagen accumulation (10). An interesting study has shown that the signaling and regulating pathways of IL-6 differ between macrophages and myocytes. It appears that unlike IL-6 signaling in macrophages, which is dependent upon activation of the NF-κB signaling pathway, intramuscular IL-6 expression is regulated by a network of signaling cascades, including the Ca2+/NFAT and glycogen/p38 MAPK pathways (11, 12). Human keratinocytes also have IL-6 receptors. We thought that the IL-6 might be an important regulator in the successful wound healing process.
Hyaluronic acid (HA) is a large glycosaminoglycan and a major extracellular component of skin, where it is involved in tissue repair. It has unique physicochemical properties and non-immunogenicity is promising for plastic surgery and therapeutic medical applications (13, 14). HA serves as an integral part of the extracellular matrix of basal keratinocytes in epidermis and involves the proliferation and migration of keratinocytes in the re-epithelialization process (12, 15). HA has crucial functions in epidermis that its free-radical scavenging functions due to and its role in keratinocyte proliferation and migration for the re-epithelialization process. In addition, studies on HA insufficient to causes increased scarring of wound healing such as keloid (1, 16). In contrast, the HA-rich fetal wound allows for scarless healing (17).
Intercellular adhesion molecule-1 (ICAM-1), CD44 and receptor for HA-mediated motility (RHAMM) were identified as major cell receptors for HA. ICAM-1 is a ligand for integrin, and CD44 can interact with collagens before their HA binding was discovered (14). The HA also contributes to tissue remodeling, proliferation and migration of cells, and participates in many interactions with cell surface receptors, notably those including its primary receptors, CD44 and RHAMM. The RHAMM also interacts with ERK to sustain basal motility, and RHAMM is necessary for CD44-midiated skin wound healing (18, 19). Activation and phosphorylation of ERK relate to cell migration during wound healing (20). It is involved in many cellular processes by interact to CD44 and ICAM-1.
The purpose of this study was to investigate the effects of IL-6 and/or HA on wound healing process in human keratinocytes cell lines (HaCaT). In this study, we firstly evaluated the combining effect of IL-6 and HA on the cell migration in scratch based wound healing assay with HaCaT cells. We further demonstrated that activation of ERK and NF-κB might involve in the cell migration of wound healing process.
MATERIALS AND METHODS
Cell culture and reagents
HaCaT cells of a human keratinocyte cell line were generously provided by Dr. JH Pak (Asan Institute for Life Sciences, Seoul, Korea) and cultured in Dulbecco's modified Eagle medium (DMEM; Invitrogen, Carlsbad, CA, USA) containing 10% fetal bovine serum (FBS; Gibco, Grand Island, NY, USA) and 1% streptomycin/penicillin (Lonza, Rockland, USA) with 5% CO2 at 37℃ incubator (Sanyo, Osaka, Japan). High molecular weight HA (Shiseido Sodium Hyaluronate, MW: 1.46 MDa, Viscosity: 2.3 M3/kg) was purchased from Shiseido Co., Ltd (Kakegawa, Japan). Recombinant human IL-6 was purchased from YbdY Biotechnology (Seoul, Korea).
MTT assay
Cell viability and proliferation was determined more than 3 times using 3-(4,5-dimethyl-2-thiazyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) reagent (Invitrogen, Carlsbad, CA, USA). Briefly, cells were placed in 96-well plates and incubated for 24 hr at 37℃ under 5% CO2, at which point 100 µL of 0.5 mg/mL MTT solution was added to each well and incubated for 4 hr at 37℃. Formazan absorbance was read at 490 nm using a GENios ELISA reader (TECAN, Mannedorf, Switzerland).
Cell-based scratch assay
The cell-based scratch assay has been previously described (21). In brief, HaCaT cells were cultured in a 6-well culture plate for 24 hr up to 90%-100% confluences of the base was filled. Scratched wound lines on the upside of cultured cells were created by 200 µL yellow micropipette tip. The scratched cells were washed with PBS after removal of culture media. And then the cells cultured for 0, 24, 48 hr with IL-6 (0.5 ng/mL) and HA (0.1 mg/mL) with 1% FBS. The wound area measured from the image taken with a microscope (Olympus, Tokyo, Japan) by Image J program (NIH, USA) at 3 different sites from each wound area of gaps. Three different sites of wound area were measured and averaged from four independent experiments.
Western blot analysis
Protein extracts were prepared from HaCaT cells using PRO-PREP™ Protein extraction solution. Bbicinchoninic acid (BCA) was used to quantify total protein amount (Pierce, Rockford, USA). The 50 µg of lysates were separated by Bolt 4%-12% Bis Tris Plus Gel (Invitrogen, Carlsbad, CA, USA) and transferred by iBlot system (Invitrogen, Carlsbad, CA, USA). Rabbit polyclonal antibody against to ERK, phosphorylated ERK, p38 and phosphorylated p38 (Cell Signaling Technology, Beverly, MA, USA) were used at a dilution of 1:1,000 for Western blot, respectively. The protein signals were visualized using LAS system (Fujifilm, Tokyo, Japan). To confirm equality of loading, membranes were reanalyzed for β-actin expression.
Immunocytochemistry
HaCaT cells were plated and cultured on 10 mm cover slips in 12-well plate. After treatment of IL-6 or/and HA for 1 hr, the treated cells were fixed in 4% of paraformaldehyde, and then permeablized in 0.1% Triton X-100 for 30 min. The cells were incubated with a primary antibody overnight at 4℃, and a secondary antibody for 2 hr at room temperature. The primary and secondary antibody against NF-κB was used (1:500; Cell signaling Beverly, USA) and anti-rabbit IgG-Alexa 594 (1:1,000; Cell signaling Beverly, USA), respectively. Nuclei were counterstained by Hoechst 33342. The stained cells were observed on a microscope (AX-70, Olympus, Tokyo, Japan), and nuclear translocation of NF-κB was analyzed and quantified using the i-solution software (IMT i-solution, BC, Canada) by comparing the intensity of NF-κB positive signal with the tetramethylrhodamine isothiocyanate (TRITC) filter set in a nucleus compared with the intensity of a entire cell. The translocation ratio of NF-κB was average of fifty independent measurements conducted in triplicate. The ratio of the nuclear translocation of NF-κB was determined using the following equation (15):
RESULTS
Effects of IL-6 and HA on the viability of HaCaT cells
To investigate whether IL-6 and HA effects on the viability of HaCaT cells, we determined the viability of various concentration of IL-6 and HA using the MTT assay (Fig. 1). There was no significant difference of cell viability in all treated concentrations of IL-6 (0.1-1.0 ng/mL) and HA (0.05-0.2 mg/mL). Thus, we used middle concentration of IL-6 (0.5 ng/mL) and HA (0.1 mg/mL) for following experiments, respectively.
Effects of IL-6 and HA on the migration of HaCaT cells
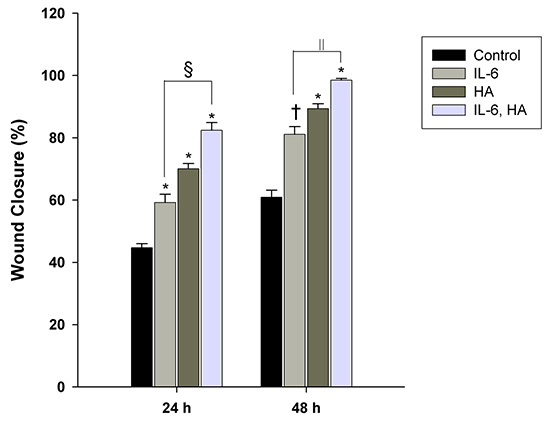
The cell-based scratch assays with HaCaT cells were performed to understand the healing mechanisms by which cell migration was contributed by IL-6 and/or HA (Fig. 2). Treatments of 0.5 ng/mL of IL-6 and 0.1 mg/mL of HA significantly increased cell migration and wound closure compared to control group (P<0.001). The wound closure of cell migration significantly increased as IL-6 (71.8%±3.1%, P<0.001) and HA (75.4%±2.6%, P<0.001) compared to control group (55.6%±1.7%) at 24 hr, and also IL-6 (82.8%±3.3%, P=0.008) and HA (91.4%±2.6%, P<0.001) compared to control group (55.6%±1.7%) at 48 hr, respectively. In addition, combining IL-6 and HA treated group synergistically enhanced the cell migration at 24 hr (81.0%± 2.7%, P=0.042) and 48 hr (95.8%±1.6%, P=0.004) compared to IL-6 group, respectively.
Activation of ERK and p38 by treatments of IL-6 and HA
To investigate the underlying mechanisms associated with IL-6 and HA, we determined the phosphorylation of ERK and p38 using the Western blot analysis. HaCaT cells were treated with IL-6 or/and HA for 1 hr and 24 hr, respectively. Phosphorylation of ERK were significantly increased by combining IL-6 and HA treated group comparing control group (P=0.009), but not IL-6 or HA for 1 hr (Fig. 3). However, there was no significant difference of p38 phosphorylation in all treated groups (Fig. 4).
Nuclear translocation of NF-κB by treatments of IL-6 and HA
Nuclear translocation of NF-κB was significantly increased in IL-6 (2.1-fold, P<0.001), HA (1.4-fold, P=0.002) and combining IL-6 and HA treated group (1.8-fold, P<0.001) compared to control group, respectively (Fig. 5). There was no synergistic effect of combining IL-6 and HA treated group in the NF-κB translocation.
DISCUSSION
Wound healing is composed of three principal phases: inflammatory, proliferative, and remodeling. The inflammatory phase initiates at the time of injury and lasts usually for 2 days. This early phase begins with reaction of hemostasis, and links to inflammatory response. Platelets form the initial thrombus and release several chemokines and growth factors. They induce the chemotaxis and activation of neutrophils and macrophages, which work together to remove bacterial debris, damaged cells and necrotic tissue from the wound. Macrophages then become the prominent cell of this phase and secret various cytokines and growth factors for alterations of cellular environment in wound. The inflammatory phase may vary in length by several factors such as infection, malnutrition, or other exogenous factors. The early stage of wound healing is related with various extracellular matrix, such as collagens, elastins, and HAs. They are regarded as a conductive environment for migration of cells into this temporary wound matrix.
Thus, we expected that IL-6 and HA treatment could promote the cell migration and activation of wound healing model. Contributions of IL-6 and HA to cell migration may attribute to its physiological functions as described above, as well as its direct interactions with cells. It have been reported that IL-6 and HA are involved in wound healing and its biological properties depend on its concentrations (7, 22, 23). We found that an underlying signaling affect associated with IL-6 and HA is synergistically activated ERK, which promotes keratinocyte migration. The observed synergistic effect of IL-6 and HA at 24 hr in HaCaT cell lines is agreeable with the understanding that wound healing processes is mediated cell migration by ERK activation (24). The ERKs are multifunctional protein kinase intracellular signaling molecules. They regulate the activation of meiosis of germ cells, mitosis of somatic cells, and differentiation of precursor cells. Various growth factors, cytokines, infections, G protein-coupled receptors, and carcinogens could activate the ERKs pathways.
Focal adhesion kinase (FAK), a non-receptor tyrosine kinase controlling cellular signaling pathways of cell migration, is in interacted with ERK pathway of the wound healing processes (20, 25). In addition, FAK/ERK-dependent pathway required for fibroblast engraftment in the decellularized mouse lung (26). In this study, the proliferation and phosphorylation of p38 MAPK were not significantly different in IL-6 and/or HA treatment, respectively. These data suggest that prompt ERK activation by combining IL-6 and HA at 1 hr would promote migration of HaCaT cells in wound healing process. We thought that enhancement of cell migration by IL-6 and/or HA may be related to the nuclear translocation of NF-κB. Taken together, these data indicate that combining of IL-6 and HA enhances wound healing by cell migration but not cell proliferation. Several observations were reported that HA interact to the CD44 and RHAMM receptors for ERK-mediated migration of the wound healing processes (27, 28, 29). The signaling pathway of this synergistic effect is not clearly defined in this study. Further studies might be required to validate the synergistic effect of IL-6 and HA on promoting cell migration in the FAK/ERK pathway and the animal models. Recently, interesting findings was reported that HA receptor of Stabilin-2 interacts with HA regulates ERK phosphorylation in arterial-venous differentiation (24).
NF-κB is one of fundamental transcription factor for cellular response and survival. It is involved in cellular responses to stimuli such as stress, cytokines, free radicals, ultraviolet irradiation, oxidized LDL, and also inflammation (30). Incorrect regulation of NF-κB has been linked to cancer, autoimmune diseases, septic shock, viral infection, and improper immune development (30, 31). In this study, we found that activation of NF-κB by nuclear translocation from cytoplasm by IL-6 and/or HA compared to control group. However, the NF-κB activation is not directly correlated to the ERK phosphorylation at 24 hr by combining IL-6 and HA treatment. We could not show the exact relationship between NF-κB and ERK in this study. It may be related to the various roles of ERK and NF-κB in different aspects of cell response and activation. Miyazaki et al. (32) showed that ERK was responsible for osteoclast survival, whereas NF-κB regulated osteoclast activation for bone resorption.
In conclusion, this is the first report that synergistic effects of combining IL-6 and HA on the cell migration of wound healing by activation of ERK and NF-κB. Thus, we suggest that combining of HA and IL-6 treatment could be used as an effective and therapeutic mixture for stimulation of cell migration in wound healing process. Further studies might be required to confirm the synergistic effects of HA and IL-6 in the animal model for the development of a novel therapeutic mixture of wound healing.



 XML Download
XML Download