

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Patients with chronic kidney disease (CKD) show evidence of increased oxidative stress, including by-products of the interactions between reactive-oxygen species (ROS) and lipids, amino acids, proteins, carbohydrates, and nucleic acids (1). Accumulating evidence shows that oxidative stress is strongly associated with increased cardiovascular morbidity and mortality in dialysis populations (2). Moreover, many inducers of CKD mediate their effects through oxidative stress, and treatment with anti-oxidants often alleviates kidney disease in humans and experimental animal models (1, 3). These findings implicate oxidative stress in the development of CKD.
The forkhead homeobox type O (FoxO) transcription factors mediate the cellular responses to oxidative stress and have been implicated in many of the ROS-regulated processes (4). FoxO transcription factors are negatively regulated by the insulin-like growth factor-1 (IGF-1) signalling pathway. Upon ligand binding, insulin or IGF-1 receptors undergo tyrosine autophosphorylation, which increases tyrosine kinase activity of the receptor and propagates signals through phosphatidylinositol 3-kinase (PI3K). Activated PI3K activates the serine/threonine protein kinase B, Akt, through the recruitment and phosphorylation of its lipid substrates (5). Akt phosphorylates and inactivates FoxO, which results in retention of FoxO in the cytoplasm and inhibition of target gene transcription. By contrast, FoxO dephosphorylation results in nuclear translocation and activation of FoxO (6). FoxOs upregulate several anti-oxidant enzymes such as manganese superoxide dismutase (MnSOD) and catalase, and protect cells from oxidative stress (7, 8). Excessive or prolonged ROS formation is implicated in a variety of human diseases, and the PI3K-Akt-FoxO signaling pathway may play a role in ROS-mediated diseases, including CKD. The pathogenic mechanisms leading to CKD result in progressive interstitial fibrosis, peritubular capillary loss with hypoxia, and destruction of functioning nephrons because of tubular atrophy (9).
To gain further insight into the mechanisms that modulate fibrogenesis of CKD, we designed the present study to investigate the hypothesis that oxidative stress promotes renal fibrosis via modulation of the PI3K-Akt-FoxO signaling pathway. We used a SOD mimetic, tempol, as an antioxidant in a mouse model of renal fibrosis, which was induced by unilateral ureteral obstruction (UUO).
MATERIALS AND METHODS
Experimental methods
Seven- or eight-week-old male C57BL/6 mice were purchased from OrientBio, Inc. (Seoul, Korea). After one week of acclimation, animals were performed UUO surgery with or without treatment with 1 mM/L of tempol (Sigma-Aldrich, Steinheim, Germany) in drinking water. Animals were divided into five experimental groups: sham, UUO-control or UUO-tempol day 3, and UUO-control or UUO-tempol day 7 (n=8, each group). Mice were sacrificed 3 or 7 days after UUO surgery, and sham-operated and obstructed kidneys were harvested.
Total collagen assay
The total collagen contents in kidney tissues were measured by acid hydrolysis of the kidney tissue section using citric acid collagen buffer (0.23 M/L citric acid, 0.88 M/L sodium acetate trihydrate, 0.85 M/L sodium hydroxide, and 1.2% acetic acid), chloramine-T solution (1.4% chloramine-T and 10% n-propanol in citric acid buffer) and Ehrlich's reagent (15% 4-dimethylamino-benzaldehyde, 62% n-propanol, and 18% perchloric acid) as described previously (10). The amount of hydroxyproline was measured by a spectrophotometric assay at 550 nm. The amount of total collagen in the kidney tissue was calculated on the assumption that collagen contains 12.7% hydroxyproline by weight.
Histological examination
Kidney sections were stained with Masson trichrome to evaluate the severity of tubulointerstitial fibrosis. Fibrotic area was quantified using MetaMorph imaging software (Molecular Devices Inc., Downingtown, PA, USA) in 10 randomly selected fields from cortex and medulla. The ratio of the fibrotic area to the total selected field was indicated as the severity of tubulointerstitial fibrosis.
Immunohistochemistry for F4/80, α-smooth muscle actin (α-SMA, Abcam, Cambridge, UK) and TdT-mediated dUTP nick end labelling (TUNEL) were performed with paraffin-embedded tissues. F4/80-positive or TUNEL-positive cells were counted in 20 views of renal cortex and corticomedullary junction.
Western blotting
The protein concentrations of frozen kidney tissues were measured using a BCA protein assay (Pierce Biotechnology, Rockford, IL, USA). For immunodetection, the blot was incubated overnight with the primary antibodies against the following proteins: MnSOD (diluted 1:2,000, Abcam, Cambridge); catalase (1:2,000, Abcam); Bax (1:500, Santa Cruz Biotechnology, Santa Cruz, CA, USA); Bcl-2 (1:100, Santa Cruz Biotechnology); transforming growth factor (TGF)-β1 (1:4,000, R&D Systems, MN, USA); PI3K (1:1,000, Abcam); Akt (1:1,000, Cell Signaling Technology, Danvers, MA, USA), phospho-Ser473 Akt (1:1,000, Cell Signaling Technology); FoxO3a (1:1,000, Cell Signaling Technology); and phospho-Ser253 FoxO3a (1:1,000, Cell Signaling Technology). The protein bands were visualized using a chemiluminescence detection system (ImageQuant LAS 4000 mini, GE Healthcare, Piscataway, NJ, USA). Band intensities were determined with Image-Pro Plus software (Media Cybernetics Inc., Rockville, MD, USA).
Measurement of lipid peroxidation and hydrogen peroxide in the kidney tissues
To evaluate the levels of lipid peroxidation in kidney tissues, malondialdehyde concentration was measured using thiobarbituric acid-trichloroacetic acid-HCl solution (0.375% thiobarbituric acid and trichloroacetic acid in 0.25 N HCl, pH 2.0) as described previously (11). The H2O2 level in kidney tissues was measured using Fox reagent (0.25 M H2SO4, 1 M sorbitol, 25 mM ferrous ammonium sulfate, and 1 mM xylenol orange in distilled water) as described previously (11).
RESULTS
Severity of renal fibrosis and total collagen content in the kidney
Masson trichrome staining demonstrated increased extracellular matrix deposition within the tubulointerstitium after UUO in a time-dependent manner. The fibrotic areas did not show significant differences between UUO-control group and UUO-tempol group on day 3, however, those in the obstructed kidney of UUO-tempol mice were significantly decreased compared to those of UUO-control mice on day 7 (P<0.05) (Fig. 1, Table 1).
Total collagen content determined by the hydroxyproline assay showed a similar pattern with the severity of renal fibrosis. The amount of total collagen was significantly lower in the obstructed kidneys from UUO-tempol mice compared with those from UUO-control mice on day 7 (P<0.05) (Table 1).
The immunohistochemistry of α-SMA, a marker of myofibroblast, demonstrated that the α-SMA-positive areas increased after UUO in a time-dependent manner. The α-SMA-positive areas in the obstructed kidney of UUO-tempol mice were significantly decreased compared to those of UUO-control mice on both day 3 and day 7 (P<0.05) (Fig. 1, Table 1).
Macrophage infiltration into the kidney
Immunohistochemistry for F4/80-positive interstitial macrophages was performed to assess the degree of interstitial inflammation (Fig. 1). The number of F4/80-positive cells significantly increased after UUO in a time-dependent manner. Tempol treatment prominently decreased the infiltration of F4/80-positive cells in the obstructed kidneys compared with control mice on both day 3 and day 7 (P<0.05) (Fig. 1, Table 1).
Renal oxidative stress
Oxidative stress in renal tissue was assessed by the expression of MnSOD and catalase by western blotting, and measuring the levels of H2O2 and lipid peroxidation. The expression of MnSOD was decreased after UUO in a time-dependent manner. Tempol treatment significantly increased the expression of MnSOD in the obstructed kidneys compared with control mice on both day 3 and day 7 (control vs tempol: day 3, 0.82±0.09 vs 1.16±0.12 fold, P=0.045; day 7, 0.23±0.04 vs 0.58±0.08 fold, P=0.04) (Fig. 2A, B). The expression of catalase, an H2O2-inducible anti-oxidant enzyme, showed no significant differences between UUO-control mice and UUO-tempol mice on day 3. It was significantly greater in the obstructed kidneys from UUO-tempol mice compared with those from UUO-control mice on day 7 (control vs tempol: day 3, 0.60±0.06 vs 0.72±0.13 fold, P=0.14; day 7, 0.31±0.05 vs 0.74±0.24 fold, P=0.02) (Fig. 2A, C).
The levels of tissue H2O2 and lipid peroxidation in the kidney also significantly increased in response to UUO. As compatible with the expression of catalase, tempol decreased the level of tissue H2O2 in the obstructed kidneys on day 7 (P<0.05) (Table 1). The level of lipid peroxidation, a critical biomarker of free radical-mediated oxidative stress, decreased in the obstructed kidneys from UUO-tempol mice compared to those from UUO-control mice on both day 3 and day 7 (P<0.05) (Table 1).
Renal apoptosis
To investigate oxidative stress-induced apoptosis, the expressions of TUNEL-positive cells, Bax and Bcl-2 and were evaluated. In TUNEL staining, UUO increased the number of TUNEL-positive cells in the obstructed kidneys. Tempol treatment markedly decreased the number of TUNEL-positive cells on both day 3 and day 7 (P<0.05) (Fig. 3A, B).
In western blotting, there was no difference in the Bax expression between the groups, however, tempol treatment increased the Bcl-2 expressions, which resulted in the relative decrease in the ratio of Bax to Bcl-2 expression in the obstructed kidneys from UUO-tempol mice compared with UUO-control mice on both day 3 and day 7 (control vs tempol: day 3, 1.28±0.04 vs 0.83±0.04 fold, P=0.03; day 7, 2.25±0.36 vs 1.21±0.21 fold, P=0.02) (Fig. 4A, B).
Expressions of TGF-β1
In western blotting, the expression of TGF-β1 in the obstructed kidneys was not different between UUO-control mice and UUO-tempol mice on day 3. However, the expression of TGF-β1 was significantly decreased in UUO-tempol group compared to UUO-control group on day 7 (control vs tempol: day 3, 4.00±0.27 vs 4.20±1.02 fold, P=0.58; day 7, 4.80±1.11 vs 3.20±0.88 fold, P=0.042) (Fig. 5A, B).
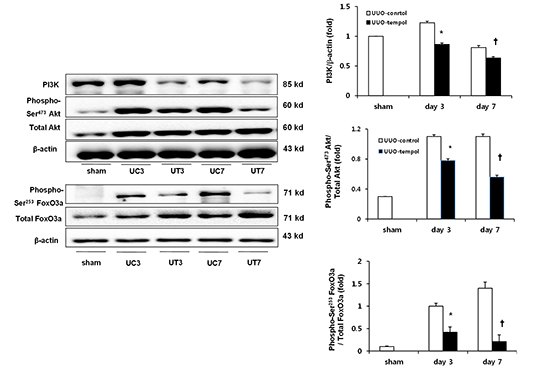
Expressions of PI3K, Akt, and FoxO3a
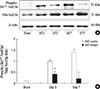
The levels of intra-renal PI3K, phosphorylated Akt, and total Akt expressions were determined using western blot analysis (Fig. 6A). The expression of PI3K was significantly lower in the obstructed kidneys from UUO-tempol mice compared with UUO-control mice on both day 3 and day 7 (control vs tempol: day 3, 1.21±0.03 vs 0.83±0.01 fold, P=0.02; day 7, 0.93±0.02 vs 0.66±0.02 fold, P=0.03) (Fig. 6B). There was no difference in the expression of total Akt between the groups of UUO with or without tempol teatment, however, the ratio of phosphorylated Akt to total Akt expression showed markedly lower values in the obstructed kidneys from UUO-tempol mice compared with UUO-control mice on both day 3 and day 7 (control vs tempol: day 3, 1.18±0.02 vs 0.79±0.02 fold, P=0.03; day 7, 1.16±0.03 vs 0.63±0.02 fold, P=0.01) (Fig. 6C).
Because PI3K activation and the phosphorylation of Akt represent negative regulators of the FoxO3a transcriptional activity, the changes of the phosphorylated FoxO3a and total FoxO3a were examined (Fig. 7A). As similar to the expressions of phosphorylated Akt and total Akt, the ratio of phosphorylated FoxO3a to total FoxO3a expression was significantly lower in the obstructed kidneys from UUO-tempol mice compared with UUO-control mice on both day 3 and 7 (control vs tempol: day 3, 0.99±0.12 vs 0.46±0.22 fold, P=0.04; day 7, 1.41±0.24 vs 0.28±0.32 fold, P=0.01) (Fig. 7B).
DISCUSSION
The purpose of the present study was to determine whether oxidative stress contributes to renal fibrosis via modulating PI3K-Akt-FoxO signaling. Tempol attenuated renal inflammation and fibrosis via inactivating PI3K and Akt and subsequently activating FoxO3a, MnSOD, and catalase. These findings suggest that therapeutic approaches to modulate PI3K-Akt-FoxO3a signaling may prevent renal damage in CKD.
Increasing evidence shows that oxidative stress is implicated in the pathogenesis of CKD and that antioxidant treatment can improve CKD (1). Tempol, a SOD mimetic, is one of antioxidants that have been shown to have renoprotective effects in animal models of CKD (12-16). However, the effect of tempol on renal fibrosis induced by UUO has not been reported. In the current study, the administration of tempol attenuated renal oxidative stress by upregulating MnSOD and catalase, and reducing the levels of ROS and lipid peroxidation. Finally, tempol reduced fibrotic areas, total collagen accumulation and interstitial infiltration of myofibroblasts and inflammatory macrophages in the obstructed kidneys. These results clearly demonstrate that tempol alleviates renal injury by increasing oxidative stress resistance and decreasing oxidative stress in mice with UUO.
Lipid peroxidation in cellular membranes is a result of ROS chain reactions of polyunsaturated fatty acids during oxidative stress and it produces several reactive aldehydes. The resultant aldehydes including malondialdehyde, a marker of lipid peroxidation, are known to induce cellular apoptosis (17). MnSOD, as well as catalase, converts superoxide anion (O2-) to the less reactive species hydrogen peroxide (H2O2). In this study, levels of malondialdehyde and H2O2 production were increased in the obstructed kidneys in a time-dependent manner and decreased with the administration of tempol, which was accompanied by increased expressions of MnSOD and catalase.
FoxO and oxidative stress are closely related. Increased cellular oxidative stress regulates FoxO post-translational modifications, and FoxO activity regulates resistance to cellular oxidative stress. The activity of FoxO is controlled by phosphorylation, acetylation, and ubiquitination (8). FoxO phosphorylation is induced by insulin/IGF-1 signaling and increased cellular stress, mainly through PI3K-mediated Akt activation (4). PI3K-Akt signaling plays a vital role in cell cycle arrest and cell survival. Phosphorylation of FoxO by Akt results in the retention of FoxO in the cytoplasm and inhibition of target gene transcription (6). FoxOs protect cells from oxidative stress by upregulating several antioxidant enzymes, such as MnSOD and catalase (7, 8). Mammals contain four distinct isoforms of FoxO: FoxO1, FoxO3a, FoxO4, and FoxO6, and FoxO3a protein is a regulator of detoxification of ROS, DNA repair, cell cycle and growth, and cellular death and survival (18). In this study, UUO prominently increased the expressions of PI3K, phosphylated Akt, and phosphorylated FoxO3a. In addition, the administration of tempol significantly decreased the renal expression of PI3K, phosphorylated Akt, and phosphorylated FoxO3a in the obstructed kidneys. These findings support that PI3K-Akt-FoxO3a signaling may be involved in renal fibrosis induced by UUO. Furthermore, greater renal expression of MnSOD and catalase was observed in the obstructed kidneys of tempol-treated mice. These findings demonstrate that tempol increases resistance to oxidative stress by modulation of PI3K-Akt-FoxO3a signaling, and subsequently upregulating MnSOD and catalase, which were known as the target molecules of FoxO3a.
In the present study, tempol also ameliorated renal apoptosis induced by UUO, which was accompanied by the activation of FoxO3a and reduction in the ratio of Bax (pro-apoptotic protein) to Bcl-2 (anti-apoptotic protein). These results are similar to our previous study showing that the decreased expression of phosphorylated FoxO3a (activation of FoxO3a) was accompanied by the increased expression of anti-apoptotic protein, Bcl-2, without change of pro-apoptotic protein, Bax (12). There are some conflicts for the effects of FoxO3a on pro-apoptotic or anti-apoptotic pathway. A previous study, contrast with the our results, showed that FoxOs regulate cell death by increasing the transcription of pro-apoptotic factors including Bcl-2 family proteins, Fas ligands, and TRAIL (19). However, another study showed that FoxO3a suppresses ROS-induced apoptosis in differentiated 3T3-L1 adipocytes, whereas FoxO3a mediates ROS-induced apoptosis in undifferentiated 3T3-L1 cells via switching to increased expression of ROS-scavenging enzymes (20). These findings suggest that the pro-apoptotic and anti-apoptotic roles of the FoxOs are highly cell and context dependent (21). Therefore, therapeutic approaches that confer gain-of-function or loss-of-function of FoxO activity may be beneficial in preventing renal fibrosis in CKD.
The present study suggests the potential interaction between tempol and PI3K-Akt-FoxO3a signalling in a UUO model, however, we did not elucidate the molecular mechanism. There are two possible hypotheses. First, tempol may activate the peroxisome proliferator-activated receptor-α (PPARα). Previously it was reported that PPARα protein expression was significantly decreased in mouse kidney after UUO, and PPARα agonist nearly normalized the expression of PPARα (22). We previously demonstrated that tempol upregulated the PPARα expression and activated FoxO3a in high-fat diet-fed spontaneously hypertensive rat kidney (12). Therefore, PPARα activation may be involved in the interaction between tempol and PI3K-Akt-FoxO3a signaling in a UUO model. Second, there is a possibility of interplay between tempol and p38 mitogen-activated protein kinase (MAPK). It was previously reported that the pre-treatment of tempol abolished ROS-mediated p38 MAPK activation and resulted in the amelioration of gastric inflammatory damage in rats under cold immobilization stress (23). Additionally, the stress-activated protein kinase p38 directly induced nuclear translocation of FoxO3a in response to doxorubicin (24). These findings suggest that tempol may regulate FoxO3a signaling by p38 MAPK in a UUO model.
In conclusion, we demonstrated that the antioxidant treatment with tempol attenuated renal fibrosis, inflammation, oxidative stress and apoptosis induced by UUO, and that the modulation of PI3K-Akt-FoxO3a signaling might be involved in this process. Because the renal fibrosis in UUO model mimics the progression of CKD in humans, our results suggest that antioxidants regulating PI3K-Akt-FoxO3a signaling may provide a therapeutic approach in the treatment of renal fibrosis in CKD.




 XML Download
XML Download