

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Prion diseases, also known as transmissible spongiform encephalopathies, are fatal neurodegenerative disorders (1). Human prion diseases, such as Creutzfeldt-Jakob disease (CJD), are characterized by the accumulation of scrapie prion protein (PrPSc) known to be protease-resistant. Pathologically, PrPSc accumulates in various types ranging from diffused deposits to amyloid plaques in different regions of the human brain (2). CJD is classified in a sporadic, familial, or acquired form (2). While the cause of sporadic CJD is yet enigmatic, familial CJD is found to be associated with specific prion protein (PrP) gene mutations (3) and acquired CJD is confirmed to be transmitted by ingestion of prion-tainted foods or various iatrogenic reasons (4).
One of the most unique features associated with prion diseases is the infectious nature of the causative agents, termed prions (5). Prion particles are composed of PrPSc, a misfolded isoform of the cellular PrP (PrPC). PrP misfolding occurs through conformational conversion of PrPC to PrPSc, in which PrPSc functions as a template (6, 7). Therefore, PrPSc can be propagated at the cells and tissues in which PrPC is expressed as a host protein. Unlike the cases of other disease-causing misfolded proteins, conversion of PrP confers propagation of PrPSc, infectivity of prions and transmissibility of diseases. Because PrPC is abundantly expressed in the central nervous system (8), the brains represent the tissue in which both PrPSc molecules and prion infectivity are found most when prions infect the hosts. Under this condition, prion infectivity is proportionally associated with the level of PrPSc molecules (9).
Because PrPSc is hydrophobic (9), it can undergo amyloidogenesis to form insoluble, misfolded PrP masses aggregated in an ordered manner (10). To date, the pathogenic role and disease transmissibility of PrP amyloids is under debate. Some argue that amyloids directly cause pathogenesis and transmission of the disease, while others propose that amyloidogenesis of PrPSc takes a role in preventing a gain-of-pathogenic function associated with non-amyloid forms of PrPSc species (11, 12). Independently, PrP amyloids are thought to be more of a neutral bystander of the pathogenic PrPSc species (13). Recent studies demonstrated that PrP amyloids play a role in propagating amyloids without causing the onset of clinical signs in animals (14). However, others claimed that PrP amyloid fibrils with the lack of any detectable PrPSc monomers were able to induce clinical and pathologic changes in hosts (15).
Although PrP amyloids are highly associated with the prognosis of prion diseases (16), much controversy exists whether PrP amyloids confer infectivity and transmissibility related to diseases. Recently, we reported generation of thioflavine T (ThT)-reactive amyloid-like PrP aggregates in oligomeric states (17). Because different assay conditions potentiate the formation of PrP amyloids with diverse structural variances, functional outcomes regarding infectivity and transmissibility of PrP aggregates generated in vitro can vary (18, 19). To address whether PrP amyloids generated with purified recombinant protein are capable of infecting hosts at the cellular level, we performed infection study of cultured cells overexpressing PrPC using PrP amyloids that we recently generated in vitro along with mouse-adapted scrapie prions. Here, we report the results of this investigation.
MATERIALS AND METHODS
Cloning for PrP genes
Full length PrP cDNA fragment encoding PrP (23-230) was amplified using an upstream primer with an EcoRI restriction enzyme recognition site and a downstream primer with a NotI restriction enzyme recognition site. PrP cDNAs previously cloned in pSP72 was used as templates. The conventional PCR procedures and conditions were used for DNA amplification. Amplified PCR products of human (Hu), mouse (Mo), and bovine (Bo) PrP were digested with EcoRI and NotI and ligated into the pIRESpuro3 vector linearized with same restriction enzymes. Then, we transformed the competent E. coli cells (DH5α) using ligation reactions. After plasmid mini-scale preparation, recombinant plasmid DNA clones were confirmed by restriction enzyme digestion and DNA sequencing.
Generation of stable cell line expressing PrPC
Stable transfection was performed as described in earlier studies with minor modifications (20). RK13, a rabbit kidney cell line, which lacks endogenous PrPC expression, was cultured in Dulbecco's Modified Eagle Medium (DMEM, high glucose) supplemented with 10% fetal bovine serum, 1% glutamax, and 1% streptomycin/penicillin under 5µ CO2 and saturated humidity conditions. RK13 cells with 50% confluency on 12-well plates (Corning, Corning, NY, USA) were transfected with 5 µg of each recombinant plasmid and empty pIRESpuro3 vector, using Lipofectamine (Invitrogen, Carlsbad, CA, USA). The stable transfectants were selected by extended cell culture in the presence of puromycine at the 100 µg/mL concentration. Individual clones of stable transfectants were established and expression of PrP genes was confirmed by genomic PCR and western blotting.
Preparation of PrP aggregate inoculum
PrP aggregates were generated in PrP amyloid formation assay (PAFA) reactions as described in our previous publication (17). Recombinant PrP (rPrP) was prepared in E. coli. The cDNA fragment encoding the C-terminal core of MoPrP (MoPrP89-230) was cloned into plasmid pET100/D-TOPO (Invitrogen). Recombinant plasmid was used for transformation of E. coli BL21 Star (DE3) (Invitrogen). Bacterial cells were grown at 37℃ until OD600=0.5 and then cultured for additional 5 hr in the presence of 1 mM IPTG. Expression of rPrP induced in bacterial cells was monitored by rPrP band detection after polyacrylamide gel electrophoresis. Harvested cells were lysed using CelLytic B Lysis reagent (Sigma-Aldrich, St. Louis, MO, USA). Inclusion bodies separated from cell lysate were then solubilized using CelLytic IB (Sigma-Aldrich). His-tag rPrP was purified using Ni-NTA agarose affinity chromatography. Purified rPrP was used to produce PrP aggregates. Briefly, 10 µg of rPrP was mixed in 0.2 mL reaction buffer (phosphate buffered saline [PBS], pH 7.0, 0.2 M guanidine hydrochloride and 10 µM ThT [Sigma-Aldrich]). Generation of PrP aggregates was monitored in situ by reading fluorescence of amyloids bound with ThT. Samples were placed in a 96-Well Black UniPlateTM Microplate (GE Healthcare, Piscataway, NJ, USA) and incubated at 37℃ for 36 hr with continuous shaking at 300 revolutions per min. To be used as an inoculum, PrP aggregates were buffer-exchanged using PBS. Five µg of PrP aggregate was used for in vitro infection of cultured cells.
Preparation of RML prion inoculum
Brain homogenates (10% w/v) collected from RML scrapie-infected and uninfected CD-1 mice were prepared in PBS as inocula (21). Mouse whole brain tissue was homogenized by serially passing multiple times through hypodermic needles from 16 to 26 gauges. After multiple rounds of freeze-and-thaw cycles, brain homogenates were centrifuged briefly at 3,000×g to separate the supernatant from tissue debris. The supernatant was stored at -80℃ until used as inocula.
Cell infection
Infection of cultured cells was performed as described in our previous publication with minor modifications (22). A series of RK13 cells (-4×103cells) cultured in 6-well plates using 3 mL of regular growing media (DMEM supplemented with 10% fetal bovine serum, 1% glutamax, and 1% streptomycin/penicillin) were infected with 100 µL of inocula. After incubation for four days, cells were cultured for several passages in the regular growing media in which inocula were no longer included.
Western blot analysis
To determine prion infectivity of RML prions or PrP aggregates, cell lysates were subjected to western blotting for proteinase K (PK)-resistant PrPSc because prions are composed of PrPSc and prion infectivity is proportionally correlated with the level of PrPSc. Cell lysates were prepared for immune-blots using lysis buffer (20 mM Tris, pH 8.0, 150 mM NaCl, 0.5% Nonidet-P 40 and 0.5% sodium deoxycholate). One mg of each cell lysate sample was incubated with 20 µg/mL PK (Invitrogen) for 1 hr at 37℃. After inactivation of PK with 1 mM phenylmethanesulfonyl fluoride, the samples were centrifuged using a table-top ultracentrifuge (Optima TLX Ultracentrifuge, Beckman-Coulter, Fullerton, CA, USA) at 100,000×g for 1 hr at 4℃. The pellets dissolved in 1×SDS-PAGE sample loading buffer (50 mM Tris-HCl, pH 6.8, 2% SDS, 10% glycerol, 1% 2-mercaptoethanol, 12.5 mM EDTA, 0.02% bromophenol blue) were separated in 12% Tris-Glycine SDS-PAGE gel. Separated proteins were transferred onto a PVDF membrane to detect the levels of PrPSc. Monoclonal primary antibodies utilized for PrP detection include anti-PrP Fab fragments D13 and R1 (23, 24), as well as other PrP monoclonal antibody 6H4 (25). Western blotting was detected using ECL Plus Detection reagents (GE Healthcare) and visualized through the G:BOX Chemi XR5 system (Syngene, Cambridge, UK).
Cell blot analysis
Direct detection of PK-resistant PrPSc from cell populations was performed by a previously reported method (26). RK13 cells overexpressing PrP of various species were grown on plastic coverslips placed in the wells of a 24-well plate. The cell infection procedure was identical to aforementioned steps. Cells on the coverslips were transferred to nitrocellulose membrane soaked in lysis buffer (10 mM Tris-HCl, pH 7.5, 150 mM NaCl, 0.5% deoxycholate and 0.5% Triton X-100). Then, the blot was incubated in lysis buffer with PK at 5 µg/mL for 1.5 hr at 37℃ with constant shaking. After inhibition of PK activity with 5 mM PMSF, PK-resistant proteins on the blot were denatured in buffer (10 mM Tris-HCl, pH 8.0 and 3 M guanidine isothiocyanate) for 10 min. Visualization of PK-resistant PrPSc for cell blots was identical to the method described for western blotting using anti-PrP antibodies and chemiluminescence.
RESULTS
Cloning of PrP gene of human, mouse and bovine in eukaryotic expression vector system
Molecular cloning of Hu, Mo, and Bo PrP genes in a mammalian expression vector, pIRES-puro3 was performed. First, PrP coding region was PCR-amplified. Agarose gel electrophoresis demonstrated that the obtained DNA fragments migrated between 600 and 700 bp maker bands (Fig. S1A), which correspond to the deduced lengths of DNA fragments. Including PrP cDNA coding region for mature form, initiation and stop codons for protein translation, and restriction enzyme recognition sites flanked to both ends of PrP genes, PCR products were expected to be 649 bp for Hu and Mo PrP genes and 673 bp for Bo PrP gene. To construct recombinant plasmids, DNA fragments were digested with restriction enzymes and ligated into linearized pIRES-puro3 for mammalian expression. The presence and corresponding lengths of the PrP inserts separated in the gel confirmed positive transformants for recombinant plasmids (Fig.S1B). The positive clones were further confirmed by PCR and DNA sequence determination (data not shown). The corresponding resultant recombinant plasmids for Hu, Mo and Bo PrP gene were termed pIRES-HuPrP, pIRES-MoPrP and pIRES-BoPrP, respectively.
Generation of stable RK13 cells expressing PrPC of three different species
Investigation for formation and propagation of PrPSc in living cells requires establishment of cell clones that express appropriate PrPC of different species. To generate stable cell lines, which express a high level of Hu, Mo, and Bo PrPC, the aforementioned recombinant plasmids pIRES-HuPrP, pIRES-MoPrP and pIRES-BoPrP were used for transfection of RK13 cells. Following selection, we obtained puromycine-resistant RK13 cells, indicating that selected cells constitutively express the PrP gene. We also confirmed the integration of the PrP gene in the genome of positive RK13 clones by performing genomic PCR using multiple positive clones (data not shown). Single cell cloning of stable transfectants resulted in establishment of >20 clones for each group of RK13 cells expressing Hu, Mo and Bo PrP.
To determine whether PrPC was stably expressed at the protein level, western blotting was performed. The clones which express the highest level of PrPC were selected from each group by western blotting (data not shown) and designated RK13-HuPrP, RK13-MoPrP, and RK13-BoPrP. Western blotting using three different antibodies, D13, R1 and 6H4 revealed that Hu PrPC and Mo PrPC was recognized by all three antibodies, but Bo PrPC was detected only by 6H4 (Fig. 1A, B, and C). D13 antibody detected Hu PrPC and Mo PrPC equally well, while detection of Mo PrPC by R1 antibody was not as efficient as detecting Hu PrPC by the same antibody (Fig. 1A and B), suggesting weaker affinity of R1 antibody to Mo PrPC epitope than to Hu PrPC. However, detection of PrPC from three different species was indistinguishable by 6H4 antibody (Fig. 1C). It appears that affinity of 6H4 to its epitope in Hu, Mo, and Bo PrP is almost identical. Thus, 6H4 antibody was used for PrP detection, hereafter.
To confirm that failure of Bo PrPC detection by D13 and R1 antibodies, as well as inefficient detection of Mo PrPC by R1 antibody, was not resulted from analyzing the samples with unequal amount of proteins, protein assay was performed for cell lysates of RK13-HuPrP, RK13-MoPrP, and RK13-BoPrP. The protein concentration were indistinguishable among samples (data not shown), suggesting that the same amount of protein samples were used for western blot analysis. This eliminates the possibility of aforementioned concerns, indicating that the observed results are solely due to antibody specificity.
Susceptibility of RK13-HuPrP, RK13-MoPrP, and RK13-BoPrP cells infected with PrP amyloids generated in vitro or natural RML prions
Our experimental goal was to investigate the formation and propagation of PrPSc in living cells which overexpress PrPC upon inoculation with PrP amyloids. Using rPrP, PAFA was performed to prepare inoculum. In situ detection of fluorescence produced from ThT-bound PrP amyloids demonstrated that macromolecular self-assembly was accomplished successfully (Fig. 2). A dramatic increase of PrP amyloid formation was detected after a 10 hr lag phase as previously reported.
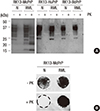
Using generated PrP amyloids, in vitro infection of RK13-HuPrP, RK13-MoPrP, and RK13-BoPrP was performed. Western blot analysis to identify de novo synthesis of PrPSc demonstrated a band pattern which resembled that of PK-resistant PrPSc at a low level from cell lysate prepared at passage 4 (Fig. 3A). Judgment based on the results of control for which RK13 cells were transfected using pIRES-puro3 with no insert, therefore expressing no PrPC, indicated that PK-resistant PrPSc-like bands did not appear to represent PrP species synthesized de novo. Instead, it could represent the PrP-Res, a form of PrP simply resistant to protease activity. Otherwise, it is likely that residual PrP amyloids from inoculum were detected. Similar analysis with cell lysate prepared at passage 8 failed to detect PK-resistant PrPSc (Fig. 3B), supporting that infection of PrPC-overexpressing cells with PrP amyloids generated in vitro did not trigger PrPSc formation and propagation.
On the other hand, infection of RK13 cells expressing PrPC of different species with mouse adapted RML prions resulted in efficient de novo synthesis of PrPSc in a species specific manner, recapitulating the species barrier effect demonstrated in animal transmission studies. RK13-MoPrP cells were susceptible to infection with RML prions, while RK13-HuPrP and RK13-BoPrP cells were not. Biochemically, PK-resistant PrPSc was detected only in RK13-MoPrP, but not in other cell lines, indicating no PrPSc propagation from Hu and Bo PrPC by mouse-adapted RML prions (Fig. 4A). Further examination of PrPSc formation in RK13-MoPrP at the cellular level showed that PK-resistant PrPSc was produced in almost all cell populations subjected to infection (Fig. 4B). These results suggest that PrPSc propagation via conformational conversion of PrPC is restricted within the same species, confirming previous observation that prion transmission is controlled by structural compatibility between PrPC and PrPSc.
DISCUSSION
To date, infectious nature of PrP amyloids is under debate (27, 28). Nonetheless, there is accumulating evidence that PrP amyloids are infectious. Abnormally folded PrP molecules can serve as seeds to accelerate amyloidogenesis in vivo and form ordered aggregates of certain suprastructure, regardless of their ability to take a role in prion pathogenesis (29, 30). Additional investigations using rPrP demonstrated that the amyloid fibrils converted from rPrP initiate prion pathogenesis in wild and transgenic mice over a long incubation period (12, 31, 32). Although the requirements of PrP amyloid formation are not necessarily addressed, lines of evidence indicated that de novo prion generated from rPrP play a role as a template to alter PrPC to PrPSc which results in clinical conditions in wild type animals (13, 33, 34).
As opposed to aforementioned observation, our results propose that PrP amyloids used for the current study lack infectious nature. To clarify the uncertainty of inherent prion infectivity of rPrP amyloids used for the study, we established cell lines that constitutively overexpress PrPC and tested whether PrPSc is synthesized de novo in the cells. Our result showed that amyloids made of recombinant MoPrP do not initiate PrPSc generation in the cells susceptible to PrPSc propagation of natural prions. This result may be explained by a few different ideas. First, the time period allowed for cells to form and propagate PrPSc during cell-based assay might be insufficient. Infection of RK13-MoPrP with RML prions resulted in detection of de novo synthesized PrPSc in cells at passage 4. However, if inherent prion infectivity of rPrP amyloids is too low, appearance of detectable level of PrPSc might require far more passages as suggested by Makarava et al. (32) who observed that prion infectivity of PrP aggregated in vitro requires a long incubation time. Second, no glycosylation on rPrP expressed in bacterial cells could explain the observed phenomenon. Generation of more pathogenic PrPSc correlates with the status of posttranslational modification (35). The PrP amyloids generated with rPrP devoid of glycosylation might have a different level of infectivity compared to natural prions. Third, the lack of lipid components in rPrP amyloids used for this study might be a cause of no or low prion infectivity. Because prion infectivity is closely associated with lipids as demonstrated in previous studies (34, 36), rPrP amyloids produced without supplementation of lipid molecules are expected to exhibit no or less prion infectivity, if any. Fourth, incongruity between PrPC expressed in RK13 cells and the misfolded conformer of rPrP amyloids might be responsible for the described observation. RK13 cells express full length PrP (23-230), while rPrP amyloids are generated with rPrP (89-230) lacking the basic N terminal region. Deletion of this region in PrP dramatically reduces formation of amyloid structure and propagation of PrPSc (17, 37, 38). Therefore, interaction of full length PrPC with truncated PrP in the misfolded states could result in inefficient PrPSc propagation. Lastly, it is also possible to speculate that the conformer found in rPrP amyloids used in this study does not represent an infectious conformer. Despite the use of identical PrP sources and experimental conditions to generate infectious synthetic prions by the same scientist group, a certain level of infectivity difference is unavoidable, presumably because formation of a certain PrP conformer, whether it is infectious or not, occurs in a random manner (11, 36). Similarly, rPrP amyloids used in this study might be composed of PrP monomers with a different conformation as compared to that of misfolded PrP species that exhibits infectivity. Therefore, conformational differences in conformers, as well as subsequent differences in supramolecular structure of rPrP amyloids and natural prion agents, could facilitate functionally different outcomes.



 XML Download
XML Download