

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Multiple system atrophy (MSA) is an adult-onset neurodegenerative disease (1) of which there are three subtypes: MSA-P (parkinsonian type), MSA-C (cerebellar dysfunction type), and MSA-A (autonomic failure type). MSA-P is the most predominant subtype (2, 3), and is characterized by severe nigral degeneration with loss of striatal neurons (4). Striatonigral degeneration (SND) is one of the pathology developing motor features in the MSA (10). As a result, it is not responsive to L-dopa (5). At present, there is no treatment available for MSA-P and the life expectancy of patients with MSA-P is lower than that of patients with Parkinson's disease (PD) (6, 7).
The first rat model of SND, which was created for the study of MSA-P, was generated by sequential injection of 6-hydroxydopamine (6-OHDA) into the medial forebrain bundle (MFB) and quinolinic acid (QA) into the striatum (8). However, this method is limited because the dopaminergic denervation that is induced by 6-OHDA affects the striatal vulnerability that is induced by the subsequent QA injection, and can therefore weaken the toxicity of QA (9, 10). A new model was generated using simultaneous unilateral injection of QA and 6-OHDA into the striatum (10). However, the loss of tyrosine hydroxylase (TH)-positive cells in the lesioned substantia nigra (SN) was only 15%, and was insufficient to cause motor deficits. It is thought that simultaneous injection of QA and 6-OHDA into the striatum allowed QA to lead a protective action against 6-OHDA-induced retrograde dopaminergic degeneration (10). These aspects were reported in previous studies (9, 11).
It is therefore necessary to develop a new model that better mimics SND. We previously showed that SND is well characterized by microPET, using the tracers 2-deoxy-2-[18F]-fluoro-D-glucose ([18F]-FDG) and [18F]-N-(3-fluoropropyl)-2-carbomethoxy-3-(4-iodophenyl) nortropane ([18F]-FP-CIT) (12). In this study, we propose a new rat model of SND that is generated by simultaneous injection of 6-OHDA into the MFB and QA into the ipsilateral striatum. We hypothesized that this method would lead to fewer adverse effects from direct or indirect interactions between the toxins, and result in a rat model of SND that closely mimics the clinical profile of this disease that is distinguishable from PD. We assessed animals using behavioral tests, multitracer microPET, and histological evaluations to explore whether this strategy creates a valid animal model of SND.
MATERIALS AND METHODS
Experimental animals
Adult male Wistar rats (Orient Bio Inc., Seongnam, Korea), weighing 250-300 g at the beginning of the experiment, were housed in a room with a 12 hr/12 hr light/dark cycle and free access to food and water. All procedures complied with the guidelines of the institutional animal care and use committee of the Asan Institute for Life Sciences, Seoul, Korea (2011-12-190).
QA and 6-OHDA lesion surgery
All surgical procedures were performed under general anesthesia induced by intraperitoneal injection of a mixture of 35 mg/kg zoletil (Virbac S.A, Carros, France) and 5 mg/kg rompun (Bayer, Leverkusen, Germany). A total of 11 rats were randomly separated into two groups. The SND group (n=6) received simultaneous injection of QA and 6-OHDA, and the PD group (n=5) received only injection of 6-OHDA. Rats in the SND group received unilateral injections of 15 µg/3 µL QA (Sigma, St. Louis, MO, USA) dissolved in phosphate-buffered saline (pH 7.4) that were targeted at the right striatum (coordinates: anteroposterior [AP], -0.4 mm; lateral [L], 4.0 mm from bregma; ventral [V], -4.0/-5.0 mm from the dura; and AP, +1.2 mm; L, 3.2 mm; V, -3.0/-4.0 mm, with the tooth bar set at -3.2 mm). A total of 40 µg/8 µL QA was delivered into all four sites at a rate of 1 µL/min using a Hamilton syringe (33-gauge) and an automated microsyringe pump (Harvard Apparatus, Holliston, MA, USA). After injection, the needle was kept in place for 3 min to prevent the solution from flowing backward, and was then retracted over the next 5 min. Following the QA injection, 8 µg 6-OHDA (Sigma) in 4 µL of 0.9% saline with 0.1% ascorbic acid was injected into the ipsilateral MFB (coordinates: AP, -2.2 mm; L, 1.5 mm; V, -8.0 mm, with the tooth bar set at +4.5 mm). The PD group received only the unilateral injection of 8 µg/4 µL 6-OHDA (Sigma) that was targeted at the right MFB and administered in the same way as for the SND group.
Stepping test and L-dopa response
The stepping test was performed six times: five times without L-dopa (during the intact state, and at 2, 4, 8, and 9 weeks post-surgery) and one time with L-dopa (at 6 weeks post-surgery). The stepping test was performed as previously described (13) with slight modifications. Briefly, both hindlimbs were firmly fixed in one hand of the experimenter, while one of the forelimbs was fixed in the other hand. The test was repeated with both the contralateral and ipsilateral forelimb. The rostral part of the rat was lowered onto a treadmill (Jeung Do Bio & Plant Co., Seoul, Korea) that was moving at rate of 0.9 m/5 sec. The rat's body remained stationary while the unilateral forelimb was allowed to spontaneously touch the moving treadmill track for >5 sec. Stepping tests were performed more than 5 sec with video recording, and the number of steps was counted during initial 5 sec. All of the experiment sessions were video recorded to allow the number of adjusted steps taken in the backward direction to be counted. Every rat performed the stepping test twice in every session, and the number of steps taken was averaged across the two trials. The number of steps taken by the contralateral forelimb was expressed as a percentage of the number taken by the ipsilateral forelimb. At the 6 week time point, 8 mg/kg L-dopa (Sigma) and 15 mg/kg benserazide (Sigma) in 0.9% saline were intraperitoneally administrated 30 min prior to each stepping test.
Apomorphine-induced rotation test
The apomorphine-induced rotation test was performed at 4 and 8 weeks post-surgery. The rotation test was performed as previously described (14) with slight modifications. Briefly, apomorphine (0.25 mg/kg, Sigma) in sterile water was subcutaneously administrated, and the rat was immediately harnessed to an automated rotometer (Panlab, Barcelona, Spain). Ipsi- and contra-lateral rotations were recorded for 45 min. The data were expressed as the net (contralateral - ipsilateral turns) average rotations per min (RPM). Rats that presented contralateral rotations <6 RPM were regarded as having SND symptoms, and rats that presented contralateral rotations >6 RPM were regarded as having PD symptoms.
PET scanning
All rats were scanned using the microPET Focus 120 system (Siemens Medical Solutions, Inc., Erlangen, Germany) with a 12×12 array of lutetium oxyorthosilicate and 96 detector blocks. All rats were assessed using [18F]-FDG to investigate metabolic activity and [18F]-FP-CIT to investigate dopamine transporter function. Scans were performed at 5 and 10 weeks post-surgery. For the [18F]-FDG PET scan, rats fasted for >12 hr and were then intravenously injected with 1 mCi [18F]-FDG in 0.2 mL of sterile saline. One hour after injection, the rats were scanned for 30 min under isoflurane anesthesia. For the [18F]-FP-CIT PET scan, rats were intravenously injected with 1 mCi [18F]-FP-CIT in 0.4 mL of sterile saline, and then immediately received dynamic scanning for 90 min. The [18F]-FDG PET data were reconstructed using a 3-dimensional (3D) maximum a posteriori (MAP) algorithm, and the [18F]-FP-CIT data were reconstructed using a 2D ordered-subset expectation maximization (OSEM 2D) algorithm (15). ASIPro software (Siemens Preclinical Solutions, Knoxville, TN, USA) was utilized to generate a three-dimensional volume of interest that was combined with three regions of interest in different axial sections. The mean uptake was quantified in the ipsilateral and the contralateral hemispheres. The data were expressed as the percentage of ipsilateral mean uptake values against contralateral mean uptake values.
Tissue processing
After the final microPET scan, the rats were transcardially perfused with 0.9% saline containing 10,000 IU heparin (Hanlim Pharm, Seoul, Korea), followed by 4% paraformaldehyde in PBS. Brains were extracted and post-fixed overnight in the same fixative, followed by dehydration in 30% sucrose until they sank. Coronal sections (40 µm thick) of the striatum (AP +2.5 to 0.0 mm) and the SN (AP -6.0 to -4.8 mm) were collected using a microtome (Leica, Wetzlar, Germany) and preserved under free-floating conditions in 0.08% sodium azide (Sigma) in PBS at 4℃. For Nissl staining, striatum sections were mounted onto gelatin-coated slides.
Histology
Immunohistochemistry was performed as previously described (16). The coronal sections of the SN (rostral section, -4.8 mm; medial section, -5.4 mm; caudal section, -6.0 mm) and striatum were selected and incubated overnight with 1:2,000 mouse anti-TH antibody (Sigma) in 0.5% bovine serum albumin in PBS (pH 7.4). They were subsequently incubated for 2 hr with biotinylated-anti mouse Ig G (1:200; Vector Laboratories, Burlingame, CA, USA) in the same solution. The complexes of primary and secondary antibody were visualized using diaminobenzidine (R&D Systems, MN, USA) colorimeric reaction. For Nissl staining, the striatum sections were rehydrated with an alcohol series of descending concentration, incubated with 0.5% cresyl violet, and dehydrated. Slides were then treated with a clearing agent and coverslipped.
Nigral cell counting and Image analysis
TH-immunopositive cells on both sides of the SN were counted in three coronal sections using a Nikon 80i microscope (Nikon, Tokyo, Japan) with NIS-Elements F3.0 software at 100× magnification. Data were expressed as the percentage of cell loss in the ipsilateral compared to the contralateral SN. The sizes of both the intact and lesioned striatum were analyzed using ImageJ 1.47t (National Institutes of Health, Bethesda, MD, USA). Data were expressed as the percentage of size in the ipsilateral compared to the contralateral striatum.
Statistical analysis
All data are presented as mean±standard deviation (SD). All statistical analyses were performed using SPSS (version 12.0.1; SPSS Inc., Chicago, IL, USA). Paired t-tests were conducted to analyze differences within groups at different time points in apomorphine test and PET data. Independent samples t-tests were conducted to analyze differences between groups at different time points in stepping, apomophine test and PET data.
RESULTS
Stepping test and L-dopa response
All rats showed motor deficit in the contralateral forelimb after lesion surgery. When the stepping test was performed with L-dopa at 6 weeks post-surgery, contralateral paw-touch rate of all rats in the PD group increased, indicating recovery to a level that was almost equal to the intact status, but L-dopa-induced dyskinesias were observed. By contrast, five of the six rats in the SND group did not show an increase in contralateral paw-touch rate or dyskinesias. There was a significant difference between the SND and PD group in the L-dopa response (P=0.003; Fig. 1).
Apomorphine-induced rotation test
Four of the six rats in the SND group exhibited bilateral touching of the cylinder wall in the apomorphine rotation test performed at 4 weeks post-surgery. The two other rats in the SND group slowly rotated in large, contralateral circles and occasionally exhibited ipsilateral turning behavior. By contrast, all five rats in the PD group showed continuous contralateral rotations. There was a significant difference in RPM between the SND and PD groups at 4 weeks post-surgery (P=0.006; Fig. 2). In the test performed 8 weeks post-surgery, two rats in the SND group showed a slight increase in contralateral rotation but the significant difference in RPM between the two groups remained (P<0.001). In the follow-up test, contralateral rotations of PD group were more increased than those in the first test (P=0.015).
MicroPET analysis
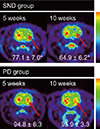
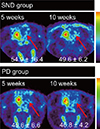
Reconstructed volumes of interest from the [18F]-FDG PET scans revealed glucose hypometabolism in the lesioned striatum of the SND group at 5 weeks post-surgery (Fig. 3). By contrast, the PD group showed bilaterally symmetrical glucose metabolism at 5 weeks post-surgery (Fig. 3). There was a significant difference in ipsilateral uptake relative to contralateral uptake between the SND group (77.1±7.0%) and the PD group (94.8±6.3%; P=0.003). This difference remained at 10 weeks post-surgery (64.9±11.4% vs. 95.9±3.3%; P=0.003). In both the SND and PD groups, [18F]-FP-CIT uptake values were lower in the lesioned striatum than in the intact striatum at 5 and 10 weeks post-surgery (Fig. 4). The relative ipsilateral uptake was similar in the SND and PD group at 5 (54.9±16.4% vs. 45.6±6.6%; P=0.246) and 10 (49.6±6.2% vs. 46.8±4.2%; P=0.441) weeks post-surgery.
Histology
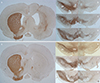
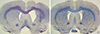
There was considerable loss of TH-immunoreactive cells in the lesioned SN in both the SND and the PD group. There was no significant difference in cell loss between the groups (74.7±19.4% in SND vs. 92.8±2.7% in PD; P=0.109; Fig. 5). Nissl staining revealed that the SND group had an enlarged ventricle and parenchymal atrophy on the lesioned side, whereas the PD group did not show differences between the two sides of the striatum (Fig. 6). The size of lesioned striatum was reduced by 48.3±12.0% compared to that of intact striatum (Fig. 5, 6).
DISCUSSION
The double toxin-double lesion strategy involving sequential injection of 6-OHDA into the MFB and QA into the striatum has been used to mimic striatonigral degeneration. The selective dopaminergic destruction induced by 6-OHDA (17) and the loss of dopamine receptors induced by QA excitotoxicity (18, 19) underlie this strategy. However, this method is not without problems. If injection of 6-OHDA into the MFB is performed first, the concentration of glutamate and aspartate in the ipsilateral striatum rapidly increases, causing down-regulation of glutamate receptors and reducing QA toxicity (9, 20, 21, 22). However, if injection of QA into the striatum is performed first, the subsequent activation of astrocytes results in a secretion of neurotrophic factors that could provide a neuroprotective effect against 6-OHDA toxicity (11). When 6-OHDA and QA were simultaneously injected into the striatum, QA reduced 6-OHDA-induced retrograde dopaminergic degeneration such that it was insufficient to generate motor deficits (10). To overcome these shortcomings, we proposed a new double toxin-double lesion strategy that involves simultaneous injection of 6-OHDA into the MFB and QA into the striatum.
The stepping test has previously been used to monitor unilateral motor deficit in the rat model of PD (23). In this study, rats that received a single injection of 6-OHDA into the MFB (PD group) showed improvement of contralateral forelimb akinesia after L-dopa administration, supporting previous results (24). On the contrary, rats that received simultaneous injection of 6-OHDA into the MFB and QA into the striatum (SND group) did not exhibit an L-dopa response because dopaminergic receptors in the ipsilateral striatum were destroyed by QA (25, 26). In addition, the PD group exhibited L-dopa-induced dyskinesias, but there were no such abnormal movements in the SND group. This mimics the L-dopa responses of humans with SND and PD.
In previous studies, the apomorphine-induced rotation test revealed an increase in contralateral rotations after injection of 6-OHDA into the MFB, and a decrease in contralateral rotations after subsequent injection of QA into the ipsilateral striatum (12, 24). However, the decrease in contralateral rotations after injection of QA into the striatum can spontaneously recover without any treatment (12, 24). The mechanism underlying this spontaneous recovery after sequential double toxin-double lesion is not clear, but it is possible that the QA-induced lesion in the striatum was insufficient to generate persistent rotation behavior (24). In this study, we observed reduced contralateral rotations at 4 and 8 weeks post-surgery in the SND group. The persistent nature of this decrease may be due to a larger lesion, as we used a different volume of toxin solution and a different concentration of QA than previous studies, or the simultaneous injection strategy. By contrast, the PD group exhibited more contralateral rotations at 8 weeks post-surgery than at 4 weeks post-surgery, possibly due to apomorphine sensitization (27). This behavioral difference between the SND and PD groups means that it will be possible to use the apomorphine-induced rotation test to evaluate the therapeutic effects of treatment in future studies.
We have introduced a multitracer microPET evaluation to clarify the exact status of SND during long term follow-up (12). MicroPET scanning is a non-invasive imaging method. Glucose hypometabolism in the lesioned striatum represents the destruction of the post-synaptic dopaminergic neuron (28). In [18F]-FDG microPET performed 5 weeks post-surgery, the SND group exhibited a lower glucose uptake in the lesioned striatum than the intact striatum, whereas the PD group exhibited relatively symmetrical glucose metabolism. This difference persisted at 10 weeks post-surgery, suggesting that [18F]-FDG microPET is a useful tool to distinguish SND from PD. In [18F]-FP-CIT microPET performed 5 weeks post-surgery, both the SND and PD group showed lower dopamine transporter activity in the lesioned striatum than in the intact striatum, indicating that this was caused by injection of 6-OHDA. This persisted at 10 weeks post-surgery, indicating that extracellular dopamine level had decreased on the lesioned side. These findings suggest that simultaneous injection of 6-OHDA into the MFB and QA into the striatum mimics the SND/MSA-P features seen in patients.
A previous study of simultaneous intrastriatal injection of 6-OHDA and QA found only a 15% loss of TH-positive cells in the lesioned SN (10). In the present study, we observed a 74.7% loss of TH-positive cells in the lesioned SN that was sufficient to induce motor deficits. This indicates that our method could produce more cell loss because we simultaneously injected both toxins into separate target areas unlikely previous method.
Nissl staining revealed enlarged ventricles and striatal atrophy on the lesioned side of the SND group. These outcomes are likely to have been caused by QA toxicity, and were not influenced by 6-OHDA.
In conclusion, we proposed a strategy of simultaneous injection of 6-OHDA into the MFB and QA into the striatum to generate a rat model of SND. Behavioral tests, multitracer microPET, and histological evaluation indicated that this strategy results in a persistent and stable rat model of SND, which may be useful for future study of SND including MSA-P.


 XML Download
XML Download