

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Cryptosporidium parvum is a protozoan parasite that causes cryptosporidiosis which is characterized by enteritis of watery diarrhea. Being a zoonotic disease, it can be transmitted between humans and animals. Numerous cryptosporidial enteritis outbreaks have been reported worldwide (1, 2, 3). In Korea, the first waterborne outbreak of cryptosporidiosis was reported in 2013 (4). Both humans and animals may be exposed to C. parvum through consumption of contaminated water and food as well as by direct contact with contaminated soil and infected hosts (5). C. parvum is released into the environment through the fecal matter of infected hosts in the form of oocysts and it remains potentially infectious for several months depending upon the environmental conditions and stresses (5). Application of manure and irrigation to agricultural fields with untreated effluent may also contribute to the prevalence of C. parvum oocysts in the farm environment (6).
Foodborne outbreaks are often not characterized as well as waterborne outbreaks (7). In the last decade, only 15 of 71 worldwide Cryptosporidium-linked outbreaks appeared to be correlated to transmission through food (8). Contaminated irrigation water appears to constitute a major route of contamination of fresh products (9). In addition, soil contaminated by manure could also be a source for contamination. There has been no report of foodborne outbreak of cryptosporidiosis in Korea until date. Additionally, little information is available regarding the contamination of soil or fresh produce by Cryptosporidium sp. in Korea. Therefore, in this study, we evaluated the extent of contamination by Cryptosporidium of both farm soil and vegetables to determine the source and route of Cryptosporidium infection.
MATERIALS AND METHODS
Sample collection and pretreatment
Soil samples were collected from 7 different localities along the western part of the Korean peninsula in January 2012 (Fig. 1). Soil (20 g) taken from each locality was mixed with 500 mL of filtered (0.22 µm) distilled water and centrifuged at 2,000×g for 20 min in an ultracentrifuge (Sorvall®Plus, Thermo Scientific, Waltham, MA, USA) after sieving through a gauze. The pellet was transferred to a 50-mL tube, washed in distilled water, and centrifuged again at 2,000×g for 20 min. The supernatant was discarded and 40 mL of distilled water was added to the pellet and mixed well. Only 1 mL of the mixed solution was taken and transferred into a microcentrifuge tube and centrifuged at 2,000×g for 10 min in a microcentrifuge (5415R, Eppendorf, Hamburg, Germany). The pellet was collected and stored at 4℃ until the DNA was purified.
Domestic food products were purchased from a local grocery market in Seoul in June 2012. Fifty grams of fresh leafy vegetables (including spinach, wild sesame leaves, and winter-grown cabbage), fruits (including blueberries, wild berries, and cherry tomatoes), and one each of root vegetables (potatoes and carrots) were put into zipper bags (Cleanwrap Inc., Gimhae, Gyeongsangnam-do, Korea). Next, 2-3 volumes of 0.1% liquinox™ (Alconox Inc., White plains, NY, USA) in 0.01 M phosphate-buffered saline (PBS; pH 7.4) was added to the bag; the bags were sealed tightly and placed on an orbital shaker (Hoefer, Holliston, MA, USA) for 15 min. The sample bags were then manually shaken repeatedly in an upside-down manner. The wash solution from the bag was collected in a bottle and centrifuged at 5,000×g for 20 min in an ultracentrifuge (Thermo Scientific). The supernatant was discarded. The food samples in the zipper bags were rewashed with 0.01 M PBS for 15 min, and the wash solution was centrifuged as before. After centrifugation, the pellets were transferred to 50-mL tubes and centrifuged at 1,000×g for 20 min in a centrifuge (Union 32R plus, Hanil Science Industrial, Incheon, Korea). Finally, the pellets were collected in microcentrifuge tubes. Cryptosporidium oocysts were isolated by using the Dynabeads® anti-Cryptosporidium kit (Invitrogen, Grand Island, NY, USA) according to the manufacturer's instructions. DNA was extracted from the soil and food samples by using the QIA-quick stool mini kit (Qiagen Inc., Valencia, CA, USA) (10).
Real-time polymerase chain reaction
The extracted DNA was used as a template in real-time polymerase chain reaction (qPCR). To obtain Cryptosporidium DNA as a positive control, C. parvum oocysts (KKU isolates) were isolated from laboratory mice (C57BL6/J) as described by Lee et al. (11). C. hominis DNA obtained from human feces (12) was used as the positive control. The primers and probes used in qPCR were designed based on the gene sequences of C. parvum SWI2/SNF2 ATPase, a Rad16 ortholog gene (Rad16, GenBank no. XM_625623.1; Table 1). Absolute quantitative qPCR reactions were performed as per the method described by Lee et al. (11). The results were analyzed using the LightCycler® software (version 4.05; Roche). DNase/RNase-free water was used instead of the template DNA as a negative control. The plasmid DNA standard for qPCR was prepared as described previously using Rad 16 as the target gene (11).
Sequences of qPCR products were confirmed using the ABI 7700 Sequence Detector and SDS software (version 1.6.3; Applied Biosystems, Foster City, CA, USA) at the Cosmo Sequence Facility Service (Seoul, Korea). The gene sequences were aligned using Clone ManagerSuite 7 (Sci-Ed Software, Cary, NC, USA). To differentiate Cryptosporidium sp. by using the restriction enzyme fragmentation method, the qPCR products were digested with TaqI restriction enzyme (Takara Bio Inc., Shiga, Japan) at 65℃ for 2 hr, and the DNA fragments were analyzed on 2% agarose gel.
RESULTS
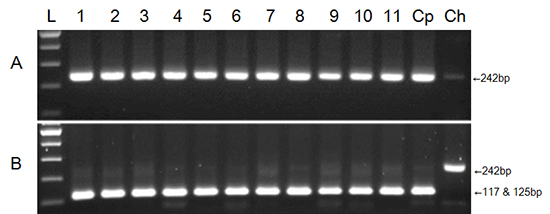
Of the 34 farm soil samples examined, Cryptosporidium sp. was detected in 11 soil samples (Table 2; 32.4% positive). The number of oocysts detected from the soil samples ranged 809-3,710/g of soil. In Hongseong-gun, Chungcheongnam-do, 6 of the 7 soil samples were positive for Cryptosporidium sp., which was the highest positive rate (85.7%) among the areas examined (except for Gyeonggi-do, where only one sample was examined). In the two locations in Jeollabuk-do, Cryptosporidium was not detected. Agarose gel electrophoresis demonstrated the presence of a 242-bp product in all Cryptosporidium-positive samples (Fig. 2A). The result of TaqI restriction enzyme digestion showed that all of the positive samples fragmented into 117- and 125-bp bands (Fig. 2B). The C. parvum DNA used as a positive control showed fragmented bands with same pattern as that of the Cryptosporidium-positive soil samples. C. hominis DNA, obtained from human diarrheal stool (12), was not fragmented by the TaqI enzyme (Fig. 2B). DNA sequence alignment data of the PCR products showed 98%-100% homology with those of C. parvum (Fig. 4A). Thus, Cryptosporidium sp. detected from the 11 soil samples were confirmed to be C. parvum.
Of the 24 food samples examined, Cryptosporidium sp. was detected from 3 food samples of carrot, winter-grown cabbage, and blueberries (12.5% positive) (Table 3). Cryptosporidium sp. was detected in only 1 of 3 samples for each kind of food examined (Table 3). The number of oocysts detected from the food samples was 42-110/g of the sample. Agarose gel electrophoresis revealed a 242-bp product from all Cryptosporidium-positive samples (Fig. 3A), and TaqI restriction enzyme digest revealed that all positive samples had fragmented into 117- and 125-bp bands (Fig. 3B). These results indicate that the food samples were contaminated with C. parvum. DNA sequence alignment data of the PCR products also supported that the Cryptosporidium sp. detected from the food samples was C. parvum (Fig. 4B).
DISCUSSION
Cryptosporidiosis is one of the most prevalent waterborne parasitic infections (1). Cryptosporidium is a zoonotic pathogen, and the sources of zoonotic contamination are usually feces-contaminated soil and water. In addition, food can mediate human infection by contamination from irrigation water and soil. The recent increase in the demand, global sourcing, and rapid transport of food products, especially of soft fruit and salad vegetables, enhances both the likelihood of surface contamination and the survival of pathogenic parasites in transmissive stages (13).
Recently, Dado et al. (14) reported that 6% of examined park soils in Madrid, Spain were contaminated with Cryptosporidium sp. and they indicated that children are mainly affected by accidental ingestion of contaminated soil. Farm soil presents an additional source of infection due to microbial pathogen contamination from agricultural runoff contaminated with livestock feces. In the present study, C. parvum was detected in 11 of the 34 (32%) farm soils examined. Animal husbandry in the vicinity of the sampling sites could be an important factor influencing the results of this study, as Cryptosporidium-positive farm areas were located near areas where animal husbandry was practiced. In addition, animal husbandry was not practiced around the farms of Jeollabuk-do, where no oocysts were detected. Cryptosporidium infection rates of cattle, goats, pigs, and livestock in some rural areas of Korea have been reported to range from 10% to 94% in 2004 and 2006 (15, 16, 17), suggesting that the source of soil contamination with Cryptosporidium oocysts could be livestock feces. Livestock studies on Cryptosporidium infection were performed in different locations in the present study, such as Jeollanam-do and Chungcheongbuk-do. In only one locality (Hongseong-gun, Chungcheongnam-do), it was reported that 1% of pigs was infected with Cryptosporidium in 2004 (15), and where 85.7% of soil samples were contaminated in 2012. This difference could be due to the difference in the soil collected from the sample site, indicating several changes in the agricultural environment in the last 10 yr in the same area.
There have been reports on foodborne cryptosporidiosis from the USA, UK, and Nordic countries (18). Cryptosporidium oocysts have been detected in several sources of fruits, salad vegetables, oysters, mussels, water spinach, and zucchini (19, 20, 21, 22, 23). Vegetables such as cilantro leaves, lettuce, carrots, and mung bean sprouts sold in the markets of Costa Rica and Norway have also been reported to be contaminated with Cryptosporidium oocysts (24, 25). In the present study, we identified Cryptosporidium sp. from carrot, winter-grown cabbage, and blueberries, and confirmed the species to be C. parvum by using a molecular genetics method. Most of the earlier studies were unable to identify the infectious Cryptosporidium sp. because of the limitations of the conventional detection systems and use of acid-fast staining or fluorescent antibody labeling techniques for confirmatory identification. Although the viability and infectivity of the detected oocysts could not be determined in this study, it is necessary to develop a washing manual for food safety practices. A previous report showed that the infective dose (ID50) for Cryptosporidium was as low as 9 oocysts (26).
Presently, there are no commonly agreed-upon guidelines to assess the methods used to detect parasitic protozoa on farm products (27). Cook et al. (28, 29) conducted extensive studies by examining a number of wash solutions and washing techniques for vegetables. The use of Alconox® for washing food samples has been reported to significantly increase the recovery efficiencies (27). Therefore, in the present study, we combined the methods described by Cook et al. (29) and Shields et al. (27) for washing the vegetables.
This study has a limitation of smaller sample size in terms of the numbers of food samples and sample sites for adequately representing the overall status of soil and food contamination in Korea. Nevertheless, this study provides valuable information about soil contamination by Cryptosporidium in Korea for the first time as well as supports the hypothesis that contaminated farm soil could be a source of contamination of farm products. Although there have been no reports of foodborne outbreaks of cryptosporidiosis in Korea yet, the results of this study indicate the potential risk of such an occurrence. Therefore, it is recommended to conduct epidemiological study on a large scale with sufficient soil and food samples on Cryptosporidium contamination in the future.




 XML Download
XML Download