

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Puberty is a complex and coordinated biologic process of sexual development that leads to complete gonadal maturation and function, and attainment of reproductive capacity. Puberty can be influenced by genetic, nutritional, environmental, and socioeconomic factors (1). The activation of pulsatile gonadotropin-releasing hormone (GnRH) secretion from specialized hypothalamic neurons to stimulate hormonal cascades and gonadal activation is a key event in the onset of puberty (2). However, the ultimate mechanisms underlying the increase in pulsatile GnRH secretion at puberty have yet to be fully elucidated (3).
Kisspeptin, the peptide product of KISS1 gene, and its putative receptor, G protein-54 (GPR54) signaling complex, have recently emerged as an essential gatekeeper of pubertal activation of GnRH neurons and the reproductive axis (4, 5, 6, 7, 8). An increase in kisspeptin signaling, which is caused by enhanced expression of KISS1 and GPR54 genes at the time of puberty, contributes to, or even drives, activation of the gonadotropic axis during pubertal development (9). Kisspeptin is a powerful stimulus for GnRH-induced gonadotropin secretion, and intermittent kisspeptin administration to immature animals was shown to induce precocious activation of the gonadotropic axis and pubertal development (10). In our previous study (11), we demonstrated that serum kisspeptin levels were significantly higher in girls with Central precocious puberty (CPP) than in their agematched pre-pubertal controls (4.61 ± 1.78 vs. 2.15 ± 1.52 pM/L, P < 0.001), consistent with the first report on kisspeptin serum levels in pre-pubertal girls and in girls with CPP in 2009 (12).
Precocious puberty is defined as the development of secondary sexual characteristics before the age of 8 yr for girls and 9 yr for boys (13). CPP is diagnosed if the process is driven by premature activation of hypothalamic GnRH secretion. As CPP may cause early epiphyseal maturation with compromised final height as well as psychological stress, early initiation of treatment is important to improve final height (14). CPP has remarkable female gender predominance and most of CPP cases are idiopathic (1, 13). However, genetic factors are known to play a fundamental role in the timing of pubertal onset (15, 16). Segregation analysis has suggested autosomal dominant transmission with incomplete, sex-dependent penetrance (15). Recently, an activating mutation of GPR54 gene was identified in a girl with CPP, implicating the kisspeptin system in the pathogenesis of sexual precocity (17). Because of the important function of kisspeptin in the regulation of puberty onset, alterations in KISS1 gene might contribute to the pathogenesis of CPP, as could alterations in GPR54 gene. Thus far, few studies, especially in Korean populations have described mutations or polymorphisms of KISS1 gene (18, 19, 20).
KISS1 gene was first discovered in 1996. This gene consists of three exons, and only parts of the second and third exons are ultimately translated into the 145-amino acid precursor peptide (21). This precursor is then cleaved into three forms of kisspeptins containing 54, 14, or 13 amino acids. The three peptides exhibit the same affinity for their single receptor since they share a common C-terminal decapeptide, designated kisspeptin-10 (5, 6).
In this study, we aimed to identify the sequence variations, including mutations and single-nucleotide polymorphisms (SNPs) of KISS1 gene. We also attempted to investigate the associations between KISS1 gene variations and CPP in Korean girls.
MATERIALS AND METHODS
Subjects
Two groups of subjects were involved in this study. Korean girls with CPP (n=143) were recruited from Korea University Ansan Hospital in Gyeonggi-do, Korea. CPP was diagnosed in accordance with the following criteria: 1) objective breast budding appearing before the age of 8 yr, 2) advanced bone age at least 1 yr ahead of the chronological age, and 3) significantly higher peak luteinizing hormone (LH) values compared with the cut-off value of 5 IU/L according to a GnRH stimulation test conducted prior to the age of 9 yr. CPP patients with identified etiology, such as a brain tumor or cranial irradiation, were excluded. The pubertal stage of each participant was determined by a pediatric endocrinologist and rated according to the Tanner criteria. Bone age, expressed in years, was determined by a single observer according to Greulich and Pyle method.
The control group (n=101) consisted of healthy Korean girls who showed breast development after the age of 9 yr. They were recruited as volunteers. The mean age of control group was 10.03±0.68 yr. The mean height and weight standard deviation scores (SDSs) were 0.66±1.16 and 1.12±1.65, respectively.
KISS1 gene analysis
Genomic DNA was isolated from the peripheral blood leukocytes of the study subjects using a DNA isolation kit (QIAamp DNA Blood Maxi prep kit; Qiagen, Valencia, CA, USA). All coding exons (exons 2 and 3) and intronic flanking regions of KISS1 gene were PCR amplified with four pairs of specific primers (Table 1). Amplification was conducted over 35 cycles, and each cycle consisted of denaturation at 95℃ for 30 sec (exon 2) or 50 sec (exon 3), annealing at 50℃ (exon 2) or 63℃ (exon 3) for 30 sec (exon 2) or 50 sec (exon 3), and extension at 72℃ for 30 sec (exon 2) or 50 sec (exon 3). An additional extension was performed at 72℃ for 10 min after the last amplification cycle. PCR was performed in a reaction volume of 20 µL containing 100 ng of genomic DNA template, 1 µM of each primer, 10 mM of each dNTP, 25 mM MgCl2, 100 mM KCl, 20 mM Tris-HCl (pH 8.3), and 1 U of Taq DNA polymerase (Takara Bio Inc., Shiga, Japan). After amplification, PCR mixtures were separated on 1.5% agarose gels with ethidium bromide to confirm the size and purity of the PCR products. Subsequently, DNA sequencing reactions were conducted using the same primer pairs and a BigDye Terminator V3.1 Cycle Sequencing kit (Applied Biosystems, Foster City, CA, USA) in accordance with the manufacturer's instructions. The sequencing reaction mixtures were electrophoresed and analyzed using an ABI3130xl Genetic Analyzer (Applied Biosystems) and Sequencing Analysis v.5.2 software.
GnRH stimulation test
The GnRH stimulation test was conducted to evaluate the pubertal status in all patients. Basal serum samples were obtained prior to GnRH (Relefact® LH-RH 0.1 mg; Aventis Pharma, Frankfurt, Germany) injection, and post-stimulation samples were acquired 30, 60, and 90 min after injection to measure LH and FSH (follicle-stimulating hormone) levels. The hormonal levels were measured using chemiluminescent microparticle immunoassays (Abbott Laboratories, Abbott Park, IL, USA).
Haplotype construction and statistical analysis
Haplotypes were estimated from genotype data for individual participants using the PHASE program, version 2.0. Deviations from Hardy-Weinberg equilibrium were assessed by comparison of observed and expected genotype frequencies with a permutation test. The IBM SPSS 20.0 Network version program was used to perform statistical analyses. Data are expressed as the mean±standard deviation (SD). Fisher's exact test and the independent t-test were used for data analysis, and P values < 0.05 were considered statistically significant. The odds ratios for SNPs were also calculated with respective 95% confidence intervals.
RESULTS
Clinical characteristics and the results of GnRH stimulation test in CPP patients
In patient group, the mean age was 8.30±0.81 yr. The mean bone age was 10.28±1.12 yr, and the mean discrepancy from the chronological age was 1.84±1.48 yr. The mean height and weight SDSs were 1.11±0.96 and 1.33±1.30, respectively. According to the results of the GnRH stimulation test, the mean basal LH levels were 0.43±0.73 IU/L, and the mean peak LH levels were 12.24±10.50 IU/L. The basal FSH levels were 2.63±1.11 IU/L, and the mean peak FSH levels were 13.77±3.91 IU/L. The peak LH/FSH ratio was 0.93±0.71. The baseline clinical characteristics and the results of GnRH stimulation tests in the patient group are summarized in Table 2.
Polymorphisms identified in KISS1 gene analysis
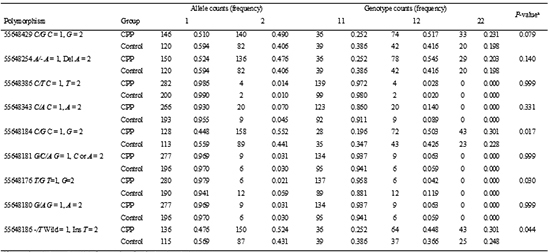
Direct sequencing of KISS1 gene revealed nine SNPs (Table 3). Among the nine polymorphisms detected in this study, 55648176 T/G was a novel polymorphism whereas the other eight have been previously reported. 55648343 C/A was previously identified in Chinese CPP patients in 2007 (19) and in Korean CPP patients in 2010 (18) and was shown to be a nonsynonymous change, leading to a substitution of P110T. Additionally, there was another nonsynonymous SNP (55648429 C/G) found that leads to the substitution of P81R. 55648386 C/T was initially identified in Korean CPP patients in 2010 (18), and was found to be synonymous. The novel SNP in this study (55648196 T/G) is located in an untranslated region. SNPs 55648184 C/G, 55648180 G/A and 55648186 -/T are also located in untranslated regions.
Comparison of allele and genotype frequencies between CPP and control groups
Allele and genotype counts and frequencies in CPP and control groups are shown in Table 4. The associations between the polymorphisms and the two phenotypes were evaluated using Fisher's exact test. No polymorphisms showed different genotype frequencies between CPP and control groups. The 55648184 C/G, 55648176 T/G, and 55648186 -/T polymorphisms showed different allele frequencies between CPP and control groups, but no significant differences in the frequencies of the other six polymorphisms were noted between the two groups.
The 55648184 C/G polymorphism was detected more frequently in CPP group than in control group (P=0.017). Using the C allele as the reference, the odds ratio of the G allele for CPP was 1.567 (95% CI, 1.091-2.252). The 55648186 -/T polymorphism was also detected more frequently in CPP group than in control group (P=0.044). Using the - allele as the reference, the odds ratio of the Ins T allele for CPP was 1.458 (95% CI, 1.015-2.095). The 55648176 T/G polymorphism was detected less frequently in CPP group than in control group (P=0.030). Using the T allele as the reference, the odds ratio of the G allele for CPP was 0.339 (95% CI, 0.125-0.920).
Comparison of clinical characteristics and the results of GnRH stimulation test between the carriers and non-carriers of gene variations in CPP subjects
Clinical characteristics and the results of GnRH stimulation test including age, height and weight SDSs, bone age, bone age advancement, basal LH and FSH levels, peak LH and FSH levels, and peak LH/FSH ratio were compared between the carriers and non-carriers of SNPs 55648184 C/G, 55648176 T/G and 55648186 -/T in CPP subjects. No parameters showed significant differences between two subgroups in CPP subjects.
Comparison of haplotype frequencies between CPP and control groups
In total, 21 haplotypes were constructed based on the typing results and their relationships with CPP were also investigated (Table 5). Haplotype GGGC-ACCC was detected less frequently in the CPP group than in the control group (P=0.024). The odds ratio of haplotype GGGC-ACCC for CPP was 0.309 (95% CI, 0.106-0.903). None of the other haplotypes differed in frequency between the two groups.
DISCUSSION
In 2003, kisspeptin was initially demonstrated to function in the reproductive axis, wherein mutations in GPR54 gene result in idiopathic hypogonadotropic hypogonadism (4, 7). These mutations suggested that kisspeptin and its receptor, GPR54, are the crucial regulators of puberty and the hypothalamus-pituitary-gonadal axis. Since 2003, further mutations have been identified in GPR54 gene and were demonstrated to cause idiopathic hypogonadotropic hypogonadism (8, 22, 23). Furthermore, an activating mutation was shown to lead to CPP (17). However, no definite causative mutation has been detected in another promising candidate gene, KISS1, in humans with idiopathic hypogonadotropic hypogonadism or CPP.
In this study, nine SNPs were detected through sequencing of KISS1 gene, and the frequencies of each SNP were calculated and compared between CPP and control groups. Although they were detected in both groups, the frequencies of SNPs 55648184 C/G and 55648186 -/T, and a novel SNP, 55648176 T/G, were found to differ significantly between the CPP and control groups. SNPs 55648184 C/G and 55648186 -/T were detected more frequently in the CPP group, and SNP 55648176 T/G was detected less frequently in the CPP group compared to the control group.
All of these SNPs are found in the untranslated region of the gene and they are all located in close proximity to each other. Untranslated regions can contain elements for controlling gene expression and it is assumed that these SNPs are located within the regulatory sequences that affect the transcription of the rest of the DNA. Changes in the DNA sequence as a result of these polymorphisms could alter the function of the regulatory sequences, subsequently altering the production of kisspeptin.
SNP 55648343 C/A was first reported in a study of Chinese CPP patients in 2007 (19), which demonstrated that p.P110T was an infrequent polymorphism with an allele frequency of 0.057 in all sequenced subjects. This allele frequency was quite similar to that in the Korean population assessed in 2010, in which the frequency for this allele was 0.046 (18). Moreover, p.P110T was found to be significantly more frequent in controls than in CPP patients in two separate studies (18, 19). It has been suggested that p.P110T may exert a protective effect on pubertal precocity (18, 19). However, these findings have yet to be confirmed by functional studies, thus more evidence is required. In the present study, the allele frequency for this polymorphism was 0.059, similar to that in the previous studies, but no significant differences in the frequencies between the two study groups were detected (P=0.331). Therefore this SNP seems not to have a critical effect on the activity of kisspeptins.
The nonsynonymous SNP 55648429 C/G was not found to be statistically associated with CPP (P=0.079). Previous association studies in Korean and Chinese populations also found no relationship between this SNP and CPP (18, 19). This polymorphism introduced a substitution of proline for arginine at the 81st, a substitution that was observed in kisspeptin-54, but not in the other three forms of kisspeptin (kisspeptin-14, -13, and -10). Therefore, the amino acid change caused by SNP 55648429 C/G would not be expected to seriously influence the bioactivity of kisspeptins. This supposition was supported by the results of a previous study showing that kisspeptin-54 was unstable and could readily degrade into kisspeptin-14, -13, and -10 (5). Thus, kisspeptin-14, -13, and -10 might be more biologically important than kisspeptin-54.
SNP 55648254 A/- leads to a frameshift mutation from the 139th site of KISS1 protein. But, there were no significant differences in the allele frequencies of this polymorphism between the CPP and control groups (P=0.140). Considering the fact that the 139th amino acid is not included in the three natural in vivo forms of kisspeptin, kisspeptin-54, -14, and -13, this frameshift mutation seems not to alter the function of kisspeptins.
A comparison of haplotypes with CPP revealed that haplotype GGGC-ACCC was detected less frequently in CPP patients than in controls (P=0.024). These findings suggest that haplotype GGGC-ACCC exerts a protective effect against CPP. Haplotype GGGC-ACCC is a combination of the G allele of SNP 55648176 T/G, the C allele of SNP 55648184 C/G, the wild-type allele of SNP 55648186 -/T, and the wild-type alleles of the other SNPs. According to the genotyping results, the G allele of SNP 55648176 T/G, C allele of SNP 55648184 C/G, and the wild-type allele of SNP 55648186 -/T were detected less frequently in the CPP group than in the control groups. Therefore, the results of the haplotype analysis suggest that the SNPs with significance do not have an effect on CPP separately, but work in combination with one another. As mentioned above, SNPs 55648176 T/G, 55648184 C/G, and 55648186 -/T are located in the untranslated region and they are in close proximity to each other. Untranslated regions can contain regulatory sequences for controlling gene expression. Accordingly, it is assumed that the SNPs identified in this study work together to affect the transcription of the rest of the DNA.
In a previous Chinese study (19), eight SNPs were detected in KISS1 gene, and only two SNPs (55648429 C/G and 55648343 C/A) identified were in common with those found in this Korean study. However, six SNPs detected in exon 3 in the current study were consistent with those identified in the previous Korean study (18). In particular, SNP 55648386 C/T, identified in the current study, was novel. The genetic background of these two ethnic groups appear to differ profoundly, even though Koreans and Chinese are both ethnically Asians.
A possible limitation of the present study was that the sample size was relatively small and P values of 0.017, 0.030, and 0.044 were not sufficiently significant. The association between gene variations and CPP could not be validated with further support from functional data. Therefore, the association should be validated by further evidence obtained from large-scaled and functional studies.
In brief, this study was aimed to identify the sequence variations and haplotypes of KISS1 gene associated with CPP in Korean girls. A novel SNP, 55648176 T/G, was identified for the first time. The allele frequencies of three SNPs, 55648184 C/G, 55648186 -/T, and 55648176 T/G, were found to differ significantly between CPP and control groups. The haplotype GGGC-ACCC was identified as one that can exert a protective effect against CPP. The genetic variations of KISS1 gene can be contributing factors of development of CPP.



 XML Download
XML Download