

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Puberty is a complex and coordinated biologic process of sexual development that lead to complete gonadal maturation and function, and attainment of reproductive capacity. The activation of pulsatile gonadotropin-releasing hormone (GnRH) secretion from specialized hypothalamic neurons to stimulate hormonal cascades and gonadal activation is a key event in the onset of puberty (1). But, the ultimate mechanisms underlying the increase in pulsatile GnRH seceretion at puberty are yet to be fully elucidated (2).
The kisspeptin, the peptide product of KISS1 gene and its putative receptor G protein coupled receptor-54 (GPR54) signaling complex has recently emerged as essential gatekeeper of pubertal activation of GnRH neurons and the reproductive axis, and it has been the focus of intense study by investigators (3-7). An increase in kisspeptin signaling which is caused by enhanced expression of KISS1 and GPR54 at the time of puberty contributes to, or even drives, the activation of the gonadotropic axis during pubertal development (8). Kisspeptin is a powerful stimulus for GnRH-induced gonadotropin secretion. Kisspeptin administration to immature animals was able to induce precocious activation of the gonadotropic axis and pubertal development (9).
Precocious puberty is defined as the development of secondary sexual characteristics before the age of 8 yr for girls and 9 yr for boys (10). Central precocious puberty (CPP) is defined if the process is driven by premature activation of hypothalamic GnRH secretion. Early diagnosis and treatment of CPP is important to improve final height because CPP can cause early epiphyseal maturation with compromised final height as well as psychological stress (11). So far, GnRH stimulation test is considered the definite biochemical parameter for diagnosis of CPP. It is highly specific but its sensitivity is low (12-14). We assumed that serum kisspeptin level may be higher in girls with CPP than in prepubertal girls because hypothalamic kisspeptin signaling increases in the onset of puberty. Up to date, no data on serum kisspeptin levels in Korean girls with CPP are available. This study was aimed to determine whether serum kisspeptin may function as a marker for CPP by investigating serum kisspeptin levels in Korean girls with CPP and healthy prepubertal controls.
MATERIALS AND METHODS
The subjects consisted of two groups : Korean girls with CPP and healthy prepubertal controls. Subjects with CPP (n = 30) were recruited from visitors to Pediatric Endocrinology Clinic at Korea University Ansan Hospital. They were between 6 and 9 yr old. They were diagnosed according to the same criteria: 1) The patients all should show Tanner breast stage scores ≥ 2 based on standardized Tanner breast stage assessment before the age of 8 yr, 2) bone age assessed by a single observer according to Greulich and Pyle method should be found to be at least 1 yr ahead of their chronological age, 3) significantly higher peak luteinizing hormone (LH) values comparing with the cut-off value of 5 IU/L under the GnRH stimulation test conducted prior to the age of 9 yr. To exclude other forms of precocious puberty, further laboratory and imaging studies were performed according to clinical judgment. CPP patients with identified etiology, such as brain tumor or cranial irradiation, were excluded. CPP patients who had been treated with GnRH agonist were excluded. The control group (n = 30) consisted of age-matched healthy prepubertal girls. They were recruited from prepubertal girls who visited the outpatient clinic of Department of Pediatrics, Korea University Ansan Hospital. They were recruited as volunteers.
Measurements of height and weight were performed in all subjects. Body mass index (BMI) was calculated as the weight in kilograms divided by the square of height in meters. The pubertal stage of each subject was determined by one pediatric endocrinologist and rated according to the Tanner criteria (15).
Blood was collected into a lavender vacutainer tube containing EDTA at 4℃ (2.5 mL of blood for each subject). The blood was transferred from the lavender vacutainer tube to the centrifuge tube. We centrifuged the blood for 15 min at 4℃ and collected the plasma. The plasma was kept at -70℃ until all samples were collected. The peptide was extracted using SEP-COLUMN (Strata, Phenomenex, Torrance, CA, USA) containing 200 mg of C18. The concentration of kisspeptin was measured using Kisspeptin-10 (Metastin [45-54]-Amide) and competitive enzyme immunoassay (Kit# EK-048-56, Phoenix Pharmaceuticals, Inc., Burlingame, CA, USA). The method was described in the product insert. Fifty-five microliters of plasma with 25 µL of primary antibody and 25 µL of biotinylated peptide were incubated at room temperature for 2 hr. Then, 0.1 mL of streptavidin-horseradish peroxidase solution was added. After incubation for 1 hr at room temperature, 0.1 mL of chemical name TMB substrate solution was added. One hour later for incubation at room temperature, the reaction was stopped with 0.1 mL of 2N HCl. After loading the immunoplate onto a microtiter plate reader, we read the absorbance at 450 nm. All measurements were in duplicates and average was calculated.
The GnRH stimulation test was conducted in all subjects. Basal serum samples were obtained prior to GnRH injection. GnRH (0.1 mg) was administered intravenously for 1 min and post-stimulation samples were taken at 30, 60, and 90 min after injection for measurements of LH and follicle-stimulating hormone (FSH) levels.
The SPSS 16.0 software package (SPSS Inc.; Chicago, IL) was used to perform statistical analyses. Data are expressed as mean ± standard deviation (SD). Independent t-test and Pearson's correlation test were used for data analysis and P values of < 0.05 were considered statistically significant.
RESULTS
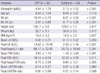
Serum kisspeptin, clinical characteristics and the results of GnRH stimulation test of CPP and control groups are shown in Table 1. Serum kisspeptin levels were significantly higher in CPP group than in control group (4.61 ± 1.78 vs 2.15 ± 1.52 pM/L, P < 0.001). CPP group also had significantly higher bone age, bone age advancement, peak LH, peak/basal LH ratio, basal FSH, basal and peak FSH, peak/basal FSH ratio and peak LH/FSH ratio compared with control group. There were no significant differences in age, height, weight, BMI, basal LH and basal LH/FSH ratio between CPP and control groups.
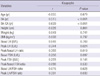
The correlations between serum kisspeptin and other clinical characteristics and the results of GnRH stimulation test are shown in Table 2. Serum kisspeptin was positively correlated with bone age, bone age advancement, peak LH, peak/basal LH ratio and peak LH/FSH ratio. No correlations were found between kisspeptin and age, height, weight, BMI, basal LH, basal and peak FSH, peak/basal FSH ratio and basal LH/FSH ratio.
DISCUSSION
To the best of our knowledge, this is the first study on serum kisspeptin levels in Korean girls with CPP or in healthy prepubertal girls. In this study, we demonstrated that serum kisspeptin level was significantly higher in girls with CPP than in their age-matched prepubertal controls. This was in accordance with the first report on the serum kisspeptin level in prepubertal girls and in girls with CPP in 2009 (16). We also found that serum kisspeptin was positively correlated with peak LH, peak/basal LH ratio and peak LH/FSH ratio during GnRH stimulation test. These findings suggest that serum kisspeptin level is associated with LH increase during GnRH stimulation test. Accordingly, serum kisspeptin may be used as an adjunctive marker for CPP because the diagnosis of CPP is based on the peak LH level during GnRH stimulation test. Moreover, it is suggested that CPP is triggered by premature increase of kisspeptin and subsequent activation of hypothalamic-pituitary-gonadal axis.
The indispensable role of kisspetin and GPR54 in the control of puberty is now well established. KISS1 gene was first discovered in 1996, and has been mapped to the long arm of chromosome 1q32. This gene consists of three exons, of which only part of the second and third exons are finally translated into a 145-amino acid precursor peptide (17). This precursor is then cleaved into three forms of kisspeptins containing 54, 14, and 13 amino acids. The three peptides exhibit the same affinity for the receptor since they share a common C-terminal decapeptide designated kisspeptin-10 (4, 7).
Kisspeptin is extraordinary potent elicitor of LH and FSH release, either after intracerebral or systemic administration of the peptide (8, 18-20). The sensitivity of LH release to the stimulatory effect of kisspeptin is very high (20, 21). Notably, FSH release in vivo appeared to be approximately 100-fold less sensitive to the stimulatory effect of kisspeptin than LH (19). Our findings that serum kisspeptin level is associated with LH increase, but not with FSH increase during GnRH stimulation test are in concordance with these results.
Gonadotropin-releasing effect of kisspeptin comes from the ability to activate GnRH release at the hypothalamus. Kisspeptin-induced membrane excitability on GnRH neurons was increased after puberty in mice (22). Activating GPR54 by kisspeptin triggers a phospholipase C-inositol triphosphate receptor-calcium cascade to initiate depolarization in GnRH neurons (23). More than 75% of GnRH neuron in the rat hypothalamus co-express GPR54 mRNA and central administration of kisspeptin induced c-fos expression (as early marker of activation) in more than 85% of GnRH neurons (24). Therefore, it has been strongly suggested that kisspeptin directly activates hypothalamic GnRH neurons, causing stimulation of GnRH secretion, which in turn elicits LH and FSH release from the pituitary (25). Notwithstanding the reports that systemic administration of kisspeptin induced LH and FSH release in animal studies (8, 26), it is not certain whether serum kisspeptin has an effect on the direct activation on hypothalamic GnRH neuron at the onset of puberty along with hypothalamic kisspeptin. Expression of KISS1 gene was demonstrated not only in brain but also in placenta, testis and small intestine (27, 28). Thus, it is assumed that there may be upstream regulatory factors of kisspeptin, which augment KISS1 gene expression not only in hypothalamus but also in other tissues at onset of puberty and consecutively serum kisspeptin level is increased. This assumption requires further investigation.
Serum kisspeptin may be used as a marker of CPP. A possible limitation of this study was that the sample size was relatively small. Therefore, our findings should be validated by large-scaled studies. Further studies on KISS1 gene polymorphisms leading to higher risk of premature increase of kisspeptin and on upstream regulator of kisspeptin are also needed.
In conclusion, serum kisspeptin level is higher in Korean girls with CPP than in age-matched prepubertal controls and CPP is supposed to be triggered by premature increase of kisspeptin.
 XML Download
XML Download