

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Differentiated thyroid cancers (DTC) maintain thyroid specific functions even with regional metastasis, respond to surgical treatment, radioiodine therapy and TSH suppression therapy. However, poorly differentiated (PDTC) or anaplastic thyroid cancers (ATC) do not respond to conventional therapy, leading to poor outcome (1). During the dedifferentiation process, the cancer loses thyroid-specific gene expression, including the ability to absorb radioiodine and to express the TSH receptor; as such radioiodine therapy or thyroid stimulating hormone (TSH) suppressive therapy is not effective in PDTC/ATC. Dedifferentiated cancers display more rapid growth properties and are refractory to chemotherapy, external beam radiation therapy, and show poor outcomes in comparison with differentiated cancers.
Redifferentiation therapy is defined as a treatment to inhibit tumor growth and regain thyroid-specific functions in PDTC and ATC as a mean of increasing the response to conventional therapy (2). The goals of such therapy are to regain the expression of differentiation markers, including the sodium iodine symporter (NIS), and to reduce levels of the dedifferentiation marker CD97. Plasma membrane expression of NIS in thyroid follicular cells is necessary to absorb iodine into the cell and, as such thyroid cancers with reduced NIS expression display impaired iodine uptake (3). CD97 is a dedifferentiation marker that is increased during cancer progression and metastasis, and is reduced by redifferentiating agent such as retinoid acid (RA) in partially dedifferentiated thyroid cancer cells (4).
Currently available redifferentiating agents include RA, histone deacetylase inhibitors (HDAC inhibitors), and peroxisome proliferator activated receptor gamma (PPARγ) agonist. Their effectiveness in dedifferentiated cancers and patient tolerability have been variable (1, 2, 5-7). Thus, this is a reason why more effective and better tolerated agents are needed.
The polyphenol phytochemicals (PPs) such as quercetin, resveratrol, genistein, and kaempferol (8-11), are present in fruits and vegetables, and inhibit cancer initiation, promotion and progression in vivo and in vitro. Moreover these compounds have been reported to induce redifferentiation in specific cancer types (12-14).
The F9 mouse embryonal carcinoma cell model (F9 cells) is frequently used to evaluate cellular morphological and molecular differentiation changes following drug and hormone treatment. The time duration of the growth cycle is very short and no self differentiation is evident; however, drugs including RA have been demonstrated to induce differentiation after 72 hr of treatment. The primitive endoderm-like structure is formed with the expression of laminin B1, collagen type IV, and tissue plasminogen activator (tPA), followed by growth inhibition (15).
In the present study, we tried to demonstrate that some PPs have the ability to induce antiproliferation and redifferentiation in thyroid cancer cell lines using the F9 embryonal carcinoma cell differentiation model.
MATERIALS AND METHODS
Cell lines and culture
F9 embryonal cancer, cells were cultured in DMEM (Sigma, St Louis, MO, USA) with 20% fetal bovine serum (FBS). TPC-1 (papillary thyroid cancer cell line) and FTC-133 (follicular thyroid cancer cell line), cells were cultured in DMEM with 10% FBS. NPA (poorly differentiated papillary thyroid cancer cell line), FRO (undifferentiated/anaplastic thyroid cancer cell line), and ARO (undifferentiated/anaplastic thyroid cancer cell line) cells were cultured in RPMI1640 (Sigma) with 10% FBS.
Reagents
Polyphenol phytochemical (resveratrol, genistein, quercetin, kaempferol, resorcinol (Sigma, Fig. 1) were dissolved in dimethylsulfoxide (DMSO; Sigma) at appropriate concentrations prior to dilution in tissue-culture medium such that the final concentration of DMSO not exceed 0.1% (v/v). Each reagent was assessed for a concentrations-dependent effect in F9 and thyroid cancer cells. In F9 cells, 0.2 µM ATRA (all-trans-retinoic acid; Sigma) was used as a positive control.
Cell proliferation and growth inhibition
Cell proliferation and growth inhibition was measured by colorimetric dimethyl-thiazol-diphenyltetrazolium bromide (MTT) proliferation assays.
RT-PCR and semi quantitative analysis
At 72 hr after the administration of reagents, RT-PCR was performed for laminin B1, collagen type IV, and tissue plasminogen activator (tPA) mRNA in F9 cell. To assess thyroid cancer redifferentiation, RT-PCR was performed for NIS, CD97 mRNA; 36B4 mRNA was assessed as a housekeeping gene. RNA extraction and complementary DNA preparation were performed by conventional methods. The primers for each gene were designed and PCRs performed according to the following conditions (annealing temperature/cycles). 36B4: forward 5'-CAGCTCTGGAGAAACTGCTG-3', reverse 5'-GTGTACTCAGTCTCCACAGA-3' (49℃/25 cycles); laminin B1: forward 5'-TGAATTCTCACTGCAGACATGGTGAAG-3', reverse 5'-ATGGATCCCTCACTTATGTCCTTAAGGA-3' (48℃/30 cycles); collagen type IV: forward 5'-ATGAATTCTCAGCGTCTGGCTTCTGCT-3', reverse: 5'-ATGGATCCGTTGCATCCTGGGATACCTG-3' (62℃/30 cycles); tPA forward 5'-TTGGGCAGAACATACAGGGTGGTC-3', reverse 5'-CTGGCATGCATCGTGGAGGTCTT-3' (62℃/30 cycles); NIS forward 5'-GCCCTCATCCTGAACCAAGTG-3', reverse 5'-TGATCCGGGAGTGGTTCTG-3' (50℃/30 cycles); CD97 forward 5'-TCCTGCCGGCAGCTCCAACC-3', reverse 5'-GGCAGCGGCAGCTGTATGAAC-3' (58℃/30 cycles). PCR was conducted for the indicated number of cycles with denaturation at 95℃ for 60 sec, annealing at the temperatures indicated for 60 sec, and extension at 72℃ for 20 sec. Each RT-PCR experiment from F9 cell and thyroid cancer cells was performed in triplicate.
In thyroid cancer cell experiments, the PCR results were analyzed by semi quantitative methods using the Total Lab TL100 program (Nonlinear Dynamics Ltd., Newcastle, UK) compensated by comparing the 36B4 density of each PCR.
Statistical analyses
Statistical analyses were performed using the SPSS software (ver. 12.0 for Windows SPSS, Chicago, IL, USA). All MTT assays were performed in triplicate and mean value were compared using Student's t-test and one-way ANOVA. A P value < 0.05 was considered to indicate statistical significance.
RESULTS
F9 cell proliferation assays
We initially assessed the effects of resorcinol, genistein, resveratrol, kaempferol and quercetin on F9 cell proliferation at concentrations of 1, 10, 50, and 100 µM respectively. Fig. 2 shows that resorcinol did not inhibit growth at concentrations below 100 µM, while genistein, resveratrol, kaempferol and quercetin all inhibited cellular growth in a dose dependent manner. Following the administration of each agent, MTT assays were performed at 24, 48, 72, 96, and 120 hr. Fig. 3 demonstrates that F9 cell growth was increased in a time-dependent manner in control cells (DMSO only), while quercetin, resveratrol and genistein inhibited cell growth up to 120 hr (P < 0.05). Kaempferol did not affect F9 cell growth.
We thus selected quercetin, resveratrol and genistein for further experiments.
Quantification of F9 cell differentiation markers by RT-PCR
Genistein, resveratrol and quercetin at concentration of 10 µM and ATRA (positive control) at a concentration of 0.2 µM were administered to F9 cells in culture. After 72 hr, RT-PCR was performed for laminin B1, tPA and collagen type IV mRNA. Fig. 4 demonstrates that genistein, resveratrol and quercetin increased the levels of these differentiation markers compared with the DMSO-alone negative control.
Thyroid cancer cell proliferation assays
Fig. 5 demonstrates that when genistein and quercetin were administered for 72 hr, the growth of TPC-1, NPA, FTC-133, FRO, and ARO cells was inhibited in a dose-dependent manner (P < 0.05). Resveratrol inhibited the growth of these cells from 10 µM excluding TPC-1 cells that displayed increased levels of growth. TPC-1 proliferation was, however, inhibited at resveratrol concentrations of 50 µM or more.
Assessment the expression of thyroid cancer differentiation markers by RT-PCR
RT-PCR for CD97 and NIS mRNA was performed following 72 hr of treatment for TPC-1 cells: genistein (100 µM), resveratrol (50 µM), quercetin (100 µM), for NPA cells: genistein (20 µM), resveratrol (50 µM), qeurcetin (100 µM), for FTC-133 cells: genistein (20 µM), resveratrol (20 µM), quercetin (20 µM), for FRO cells: genistein (50 µM), resveratrol (50 µM), quercetin (20 µM), for ARO cells: genistein (50 µM), resveratrol (20 µM), quercetin (20 µM). In contrast to the other cell lines under investigation, TPC-1 cells did not express the dediffenetiation marker CD97. We observed a decrease in CD97 expression following genistein treatment in NPA cells. Resveratrol decreased CD97 in FTC-133, NPA and marginally decreased CD97 in FRO cells, while qurcetin decreased CD97 levels in FTC-133, NPA, FRO, and ARO cells. Expression of CD97 in TPC-1 cells was unaffected by the presence of any of the PPs (Fig. 6). NIS was expressed in TPC-1 and FTC-133 cells. While the levels of NIS were unchanged in the presence of genistein, resveratrol and quercetin in TPC-1 cells, all three compounds increased NIS levels in FTC-133 cells. Genistein and resveratrol both failed to induce in NPA, FRO or ARO cells. Qurcetin did not induce NIS in ARO cells but did induce NIS in dedifferentiated/anaplastic NPA and FRO cells (Fig. 7).
DISCUSSION
RA, HDAC inhibitor, and PPARγ agonists are effective redifferentiation therapies in partially dedifferentiated thyroid cancers, but are ineffective undifferentiated/anaplastic cancers and display high patient toxicity or effectiveness only in vitro. Polyphenol phytochemicals have been reported to inhibit cancer initiation, promotion and progression in vivo or in vitro and moreover have been reported to induce redifferentiation in specific cancer types (12-14). We present the first report that identifies the ability of PPs to promote tumor growth inhibition in both partially dedifferentiated cancer and dedifferentiated/anaplastic cancers.
We demonstrated that genistein (rich in soybeans), resveratrol (rich in grapes), quercetin (rich in onions) inhibit the tumor growth of TPC-1, FTC-133, NPA, FRO, and ARO cell lines. The mechanism of the growth inhibition of genistein, resveratrol and quercetin is thought to involve the inhibition of epidermal growth factor (EGF), NF-κB, cyclin D and protein tyrosine kinase, all of which increase tumor promotion, progression, and inreases caspase which can induce apoptosis (16-18). They have been tested in normal and cancer cultured cells, and shown that these compounds induced apoptosis in cultured cancer cells but not in their normal counterparts up to 100 µM, so these effects were not by general toxicity (19). Other studies have suggested that PPs act as phytoestrogens and inhibit the estrogen-dependent growth of estrogen receptor (ER) - positive cancer cells (20) and specific cancer cells (NPA, FRO) that express ER (11). We observed increased TPC-1 cell growth in response to low dose resveratrol (10 µM), most likely caused by its phytoestrogenic effects, stimulating the estrogen receptor α (ERα) and inhibitory estrogen receptor β (ERβ). The dose of resveratrol displays biphasic effects in MCF-7 breast cancer cells and low dose are thought to affect growth stimulation (21). However, no report has described the expression of ER in TPC-1 cells.
CD97 is a representative dedifferentiation marker in thyroid cancers that is stimulated by EGF, and can be inhibited genistein, resveratrol and quercetin (16), consistent with their effects on cancer cell observed in our study.
The dedifferentiation process is accompanied by both genetic mutations and epigenetic changes. During dedifferentiation process, a loss of NIS gene expression occurs, caused by hypermethylation of the NIS gene; this is recovered by demethylating agent 5-azacytidine (22). Additionally, NIS expression is induced by TSH mediated by TSH receptor (TSHR). Dedifferentiated thyroid cancers are accompanied by a loss of the TSHR gene promoter hypermethylation and is restored by 5-azacytidine (23). DNA methylation is mediated by DNA methyltransferases (DNMT) and numerous cancers including thyroid cancers, have been reported to have stimulated hypermethylation (22, 23). In this study, genistein, resveratrol and quercetin all incresed NIS expression in FTC-133 cells while quercetin also incresed NIS expression in dedifferentiated/anaplastic NPA and FRO cells. Quercetin has been reported to reverse DNA hypermethylation in a dose-dependent manner in other cancers (24). Genistein has been reported to reverse retinoic acid receptor β (RARβ) promoter hypermethylation and increase RARβ in prostate and esophageal cancers (25). There is no report regarding the effects of resveratrol in relation to DNA methylation; however, most of PPs are substrates of catechol O-methyltransferase (COMT) and are converted to S-adenosyl-L-homocysteine (SAH) by COMT, an antagonist of DMNT (24). Additionally, resveratrol has been reported to increase the transcription of PPARγ, which can induce thyroid cancer redifferentiation (26). RA acts through binding with RARβ and has been reported to restore NIS expression in FTC-133 (27), but not in NPA, FRO or ARO cells. Additionaly RA does not inhibit growth of FRO and ARO cells that have lost RARβ (28), which occurs as a result of promoter hypermethylation in numerous thyroid cancer cell lines (29). Thus, PPs that inhibit RARβ hypermethylation, when administered in combination with low-dose RA, may be effective in thyroid cancers that are refractory to RA therapy. Serum concentrations of PPs typically do not exceed 10 µM in humans, a concentration that is both antiproliferative and induces redifferentiative in vitro. The tissue concentration levels of these compounds can increase 1-10 fold following dietary supplementation (30).
In conclusion these findings suggest that genistein, resveratrol and quercetin may be effective for thyroid cancer redifferentiation therapy, including of undifferentiated/anaplastic cancers. Further investigations are required to increase the tissue concentrations of these polyphenol phytochemicals in thyroid cancer patients and to assess their effectiveness in patients with dedifferentiated thyroid cancers.
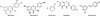





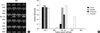
 XML Download
XML Download