

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The human paraoxonase (PON) multigene cluster is located on the long arm of chromosome 7 at q21-q22 and includes 3 members-PON1, PON2, and PON3-adjacent to each other (1). PON1 is a high-density lipoprotein (HDL)-associated serum enzyme composed of 354 amino acids (43 kDa) and the most abundant PON enzyme. PON1 and PON3 are predominantly expressed in the liver and secreted into blood (1-3). PON2 is more widely expressed in a number of tissues including the brain, liver, kidney, and testis but not detectable in the blood (4). Among them PON1 only has paraoxonase and arylesterase activities, and PON2 and PON3 lack these activities (3). It is well known that PON1 hydrolyzes the oxons (e.g., paraoxon, diazoxon, and chlorpyrifos oxon) which are toxic metabolites of organophosphate pesticides. In addition, PON1 plays an antioxidant role in lipid metabolism by inhibiting the oxidation of low-density lipoprotein (LDL) and preventing lipid peroxide accumulation on LDL (4-6). These actions of PON1 exert a protective effect on the early progression of cardiovascular disease and atherosclerosis (2).
About 200 single nucleotide polymorphisms (SNPs) have been identified in the human PON1 gene. These are located in the promoter (7 SNPs), intronic (171 SNPs), exonic (5 SNPs), and 3'-untranslated regions (15 SNPs) (7). Most studies have focused on the associations between genetic polymorphisms in the PON1 promoter region (-108CT and -162AG) and coding region (55LM and 192QR) and on PON1's catalytic activity, which is associated with susceptibility to certain pesticides and cardiovascular diseases (2-6). The PON1-108CT polymorphism in the promoter region is in linkage disequilibrium (LD) with the 55LM polymorphism and is related to variation in plasma PON1 expression levels (8). The PON1 192QR genetic polymorphism has been shown to produce a substrate-dependent difference on the catalytic efficiency of PON1. In in vitro studies, the PON1 192 R allele hydrolyzes paraoxon more efficiently than the Q allele, whereas diazoxon, sarin, and soman are hydrolyzed more rapidly by the Q allele (9). In the 3' end region of intron 5, adjacent to the PON1 192QR polymorphic site, 2 intronic polymorphisms exist at position 17899 (17899insdelTT, SNP ID: rs3917539) and 17974 (17974CT, SNP ID: rs3917541) of the PON1 gene. To date, intronic genetic polymorphisms of PON1 have not been characterized, although they are suspected of being involved in the regulation of enzyme activity through interfering with the correct splicing of the gene's mRNA (3).
PON1 phenotype can be measured by a two-substrate assay that uses the substrate pair-paraoxon and phenylacetate (10). Individual PON1 paraoxonase and arylesterase activities have been shown to be affected by genetic polymorphisms of PON1 and various environmental factors (2).
In this study, we analyzed the frequencies of 3 PON1 genetic polymorphisms in intronic (17899insdelTT and 17974CT) and exonic (192QR) regions, and evaluated the influence of intronic and exonic genetic polymorphisms of PON1 on serum enzyme activity in a Korean population.
MATERIALS AND METHODS
Study subjects
Subjects of this study consisted of 153 healthy individuals who received routine medical examinations at Chungbuk National University Hospital, Korea. Trained interviewers collected information on demographic factors, smoking habits, alcohol consumption, and occupational history, using a structured questionnaire. Peripheral blood was collected in both an EDTA-treated tube and a serum separator tube for DNA preparation and the PON1 activity assay, respectively.
PON1 genotyping
Genomic DNA was isolated from peripheral blood using a DNA Extractor WB Kit (Wako, Osaka, Japan) according to the manufacturer's protocol. The region of PON1 including the intronic (17899insdelTT and 17974CT) and exonic (192QR) SNP sites was amplified using primers 5'-TGT TAC AGT GCT ATA ATC ACC TCC-3' and 5'-CCA AGA TAT CTC CTG AGA ATC TGA-3' in a thermal cycler (TaKaRa, Shiga, Japan). The PCR were performed in 25 µL PCR reaction solution containing 50 ng of genomic DNA, 10 × PCR buffer (Roche Applied Science, Indianapolis, IN, USA), 5 pM of each primer, 200 µM each dNTP and 0.025 unit FastStart Taq DNA polymerase (Roche Applied Science). Thermal cycling was carried out as follows: 5 min of denaturation at 94℃, then 35 cycles consisting of denaturation at 94℃ for 30 sec, annealing at 52℃ for 30 sec, and extension at 72℃ for 45 sec. Unincorporated dNTPs and primers in the PCR product (484 bp fragments) were purified with a purification kit (Real Biotech Corporation, Banqiao, Taiwan). Intronic and exonic SNPs of PON1 were determined by a fluorescent dye terminator method using an ABI PRISM BigDye Terminator Cycle Sequencing Ready Reaction Kit and an ABI 3730XL capillary DNA Sequencer (Applied Biosystems, Carlsbad, CA, USA).
PON1 enzyme activity assays
Serum PON1 enzyme activity was determined by a spectrophotometeric method using 2 substrates-paraoxon and phenylacetate (10). For measurement of PON1-paraoxonase activity, 190 µL of the substrate (1.2 mM paraoxon, 0.26 mM Tris-HCl at pH 8.5, 25 mM CaCl2, and 500 mM NaCl) was mixed with 10 µL of diluted serum. Enzyme activity was calculated using the rate of p-nitrophenol production monitored at 405 nm and was expressed in units/mL on the basis of the molar extinction coefficient of 18/mM/cm for p-nitrophenol.
PON1-arylesterase activity was measured in 190 µL of the substrate (3.26 mM phenyl acetate, 9 mM Tris-HCl at pH 8 and 0.9 mM CaCl2) mixed with 10 µL of diluted serum. Enzyme activity was calculated using the rate of phenol production monitored at 270 nm and was expressed in units/mL based on the molar extinction coefficient of 1.31/mM/cm for phenol. Both enzyme activity assays were carried out in a SPECTRAmax PLUS microplate spectrophotometer (Molecular Devices, Sunnyvale, CA, USA) as described previously (11). The assay value was corrected for path-length using software provided with the microplate reader (12).
Statistical analysis
Means of serum PON1 activity according to different genotypes were compared using analysis of variance (ANOVA) and t-tests. To test the effects of 3 genetic polymorphisms of PON1 on serum PON1-paraoxonase or PON1-arylesterase activity, multiple regression models (stepwise selection) that included independent variables (e.g. gender, age, smoking, alcohol consumption, triglyceride level, HDL-cholesterol level, LDL-cholesterol level, and body mass index) were used. LD statistics (D' and chi-square) for 3 genetic polymorphisms of PON1 were determined by using the SNP Analyzer V2.0 (ISTECH Inc., Goyang, Korea). All statistical analysis was performed using SPSS 12.0 for Windows (SPSS company, Chicago, IL, USA).
RESULTS
General characteristics, lipid profile, and serum PON1 activity of study subjects
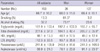
The general characteristics, lipid profile, and serum PON1 activity of study subjects are presented in Table 1. The mean age of study subjects was 66.7 ± 10.2 yr, 75% of all subjects were aged 60 yr or older. The proportion of smokers and alcohol drinkers was significantly greater in men than in women, and LDL levels were significantly higher in women than in men. The means of serum PON1-paraoxonase and arylesterase activity were 241.8 ± 105.9 unit/L and 88.9 ± 24.0 unit/mL, respectively. The serum PON1-arylesterase activity of women was significantly higher than that of men (P < 0.05) (Table 1).
Distribution of genotypes and alleles, and linkage disequilibrium analysis for PON1 SNPs
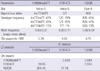
The distributions of the genotypes and alleles for PON1 intronic (17899insdelTT and 17974CT) and exonic (192QR) polymorphisms are presented in Table 2. All genotypes were in agreement with Hardy-Weinberg equilibrium in this study (P > 0.05). The frequencies of major alleles of intronic (17899insTT and 17974C) and exonic (192R) polymorphisms were 63%, 89%, and 66%, respectively. We observed significant LD between all tested SNPs, with the strongest LD found between SNPs 17899insdelTT and 192QR (D' = 0.984) (Table 2).
Effects of PON1 genetic polymorphisms on PON1 serum enzyme activity
Both the intronic and the exonic genetic polymorphisms were associated with significant differences in serum PON1-paraoxonase activity. Serum PON1-paraoxonase activity was highest for the 17899insTT/insTT, 17974C/T or T/T, and 192R/R genotypes and lowest for the 17899delTT/delTT, 17974C/C, and 192Q/Q genotypes. The mean serum PON1-arylesterase activity was highest in individuals with the 17899delTT/delTT, 17974C/T or T/T, and 192Q/Q genotypes, respectively. The 17899insdelTT and 192QR polymorphisms were associated with significant differences in serum PON1-arylesterase activity (Table 3).
Multiple regression analysis (stepwise selection) was used to assess determinants of serum PON1 activity (Table 4). Age was negatively correlated with serum PON1-paraoxonase activity with borderline significance (P = 0.091), and ever-smokers had lower serum PON1-paraoxonase activity than never-smokers (P = 0.005). Triglyceride and HDL-cholesterol were positively correlated with serum PON1-paraoxonase activity. Both intronic (17974CT) and exonic (192QR) polymorphisms were strongly associated with serum PON1-paraoxonase activity. Age, smoking history, triglyceride and HDL concentrations, body mass index, and intronic (17899insdelTT and 17974CT) and exonic (192QR) polymorphisms explained 65.8% of the variance of serum PON1-paraoxonase activity in this study (P < 0.001). The effect size of the 192QR polymorphism for serum PON1-paraoxonase activity was about 53.7% and had the largest effect of any of the independent variables. PON1-arylesterase activity was positively associated with triglyceride and HDL-cholesterol levels (P = 0.048 and 0.001, respectively) and negatively associated with age and the 192QR polymorphism in multiple regression models with serum PON1-arylesterase activity as the dependent variable. These 4 variables explained 32.1% of the variance of serum PON1-arylesterase activity. The 192QR polymorphism accounted for 20.2% of the variance of PON1-arylesterase activity and showed the largest effect of any the independent variables (Table 4).
DISCUSSION
In this study, serum PON1-arylesterase activity was higher in women than men. This gender difference was owing to the lower rate of smoking among women compared with men, rather than being due to a gender effect. As with previous epidemiologic studies (13, 14), our data suggest that smoking decreases serum PON1 activity (Tables 1, 4). It concurs with the results of an in vitro study in which cigarette smoke extracts inhibited serum PON1 activity through modification of PON's free thiol residue at amino acid 283 (15).
The frequency of the 192R allele (66%) in this study was similar to that of earlier reports on Korean (60%-66%) (16, 17), Japanese (64%-67%) (18, 19), Chinese (65%) (20), and African-American (63%-85%) populations (21, 22), but differed when compared to south-western Korean (30%) (23), Caucasians (27%-31%) (21, 24) and Hispanic (41%-46%) populations (21, 25).
As far as we know LD among intronic and exonic polymorphisms of the PON1 gene has not been studied till now. Previous studies have reported LD within the SNPs of the coding region (55ML and 192QR) or 5' flanking region (-108CT, -126GC, -162AG, -832GA, and -909CG) of PON1 (2, 8). Although our data showed LD among tested SNPs, it is difficult to exclude the possibility that LD was caused by the relatively short physical distance between the tested SNPs. Since strong LD exists between the two-base deletion/insertion polymorphism (17899insdelTT) and 192QR, which is one of the most important SNPs determining PON1 enzyme activity, primers for 192QR genotyping should not include the 2-base deletion/insertion polymorphism site.
As in previous studies, the results of this study suggest that the 192R/R genotype is associated with higher serum paraoxonase activity and lower arylesterase activity (2, 3, 9). However, the level of serum PON1 activity in our study is not directly comparable with those of previously reported data, because of various differences in environmental factors (e.g., diet, smoking, alcohol consumption, age, and environmental toxin exposure). In addition, individual HDL level can influence the serum activity of PON1, as can the 192QR genetic polymorphism (3, 9).
In this study, we first characterized the relationship between intronic polymorphisms of PON1 and serum PON1 activity. Our results demonstrated that intronic genetic polymorphisms of PON1 can also affect serum PON1 activity. Although this effect was probably due to LD of the intronic polymorphisms with the 192QR polymorphism, it cannot be ruled out that intronic SNPs of PON1 may directly regulate transcription by alteration of RNA elongation, splicing, or maturation (26-28).
The effect size of the 192QR polymorphism for serum PON1-paraoxonase activity in this study (53.7%) is similar to those of American studies, 48%-59% (25, 29). Our data showed that the activity of both serum paraoxonase and arylesterase correlated with serum HDL level. Previous studies have reported that HDL is an important determinant of serum PON1 enzyme concentration and activity, is related to the release mechanism, and acts as the serum transport vector for PON1 (10). Up to about 70% of the variance of serum HDL levels is regulated by genetic factors such as ApoA1, Abca1, Scarb1, cholesterol ester transfer protein (Cetp), and endothelial lipase (Lipg) (30). Further study is necessary to elucidate the other genetic factors associated with serum PON1 level and activity.
In conclusion, it is suggested that although the PON1 192QR SNP is primarily associated with serum PON1 activity, the intronic SNPs (17899insdelTT and 17974CT) of PON1 are also involved in serum PON1 activity, which may be mediated by LD among SNPs.


 XML Download
XML Download