

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
In the last decade, clinical observations and controlled studies have shown that leptin produced by fat tissue regulates bone metabolism through a central pathway consisting of the hypothalamus, sympathetic nervous system, and multiple intermediary steps. This relationship between fat and bone, initially thought to be unilateral, is now considered to be bilateral or reciprocal after the discovery of the role of bone in glucose and fat metabolism. This newly identified feedback loop between bone and energy metabolism is mediated by osteocalcin (OC), an osteoblast-produced protein, and has advanced academic progress in osteology and endocrinology. Furthermore, studies of OC have provided the fundamental basis of therapeutic strategies for metabolic disorders. In this review, we discuss the feedback loop between bones and fat, and new discoveries related to novel metabolic roles of OC.
ROLE OF LEPTIN IN REGULATING BONE METABOLISM
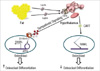
The prevailing model in skeletal biology is that bones constantly undergo the physiological process of bone remodeling, which comprises two phases: bone resorption by osteoclasts followed by bone formation by osteoblasts (1). Imbalance between the two phases whereby osteoclast activity surpasses osteoblast activity leads to decreased bone mineral density and, in turn, an increased risk of osteoporosis. A number of published studies have demonstrated that differentiation and functions of these bone-specific cells are regulated by leptin, an adipocyte-derived hormone that regulates energy intake and expenditure. Numerous other studies have examined the relationship between serum leptin levels and bone anabolism, and have proposed a bimodal response in which moderate increases in leptin stimulate bone formation whereas higher levels actually inhibit it (2-4). This bimodal response is further complicated by the finding that adipocytes in both the periphery and in the bone marrow secrete leptin, which may induce apoptosis of bone marrow stromal cells and favors bone resorption at high local concentrations (5, 6). It has been proposed that leptin exerts osteogenic functions through a peripheral pathway that directly targets a leptin-specific receptor on osteoblasts, but the results of relevant studies were inconsistent (7, 8). Other researchers have consistently shown that leptin regulates bone formation through a central pathway comprising the hypothalamus and central nervous system (9-15). Leptin is known to initiate intracellular signals within the hypothalamus through its binding to Ob-Rb, the leptin receptor isoform present in hypothalamic nuclei (16). At least two different central hypothalamic pathways, through which leptin influences bone formation in an antagonistic manner, have been identified (Fig. 1) (9-15). The first, anti-osteogenic, influence of leptin involves up-regulation of receptor activators of NF-kappa B ligand (RANKL), an osteoclast differentiation factor, through sympathetic signaling via β2-adrenergic receptors, the only adrenergic receptor present on osteoblasts (12, 15). Sympathetic signaling on β2-adrenergic receptors induces phosphorylation of activating transcription factor 4 (ATF4), a cell-specific cAMP response element binding (CREB)-related transcription factor that is essential for osteoblast differentiation and function (16). The second, more recently defined, osteogenic influence of leptin involves modulation of cocaine and amphetamine-regulated transcript (CART) (12), a hypothalamic neuropeptide encoded by the CARTPT gene whose expression is increased by leptin (17). Down-regulation of RANKL expression by CART is unfavorable to bone resorption and inhibits osteoclast differentiation (12, 13).
IMPLICATIONS OF A RECIPROCAL RELATIONSHIP BETWEEN BONE AND ENERGY METABOLISM
The concept of hormonal regulation in which hormones are regulated by a feedback loop, the facts that osteoblasts and adipocytes develop from a common precursor cell, the mesenchymal stem cell (15), and that bone remodeling is regulated by an adipocyte-derived hormone, leptin, have all aroused interest among integrative physiologists regarding the possible endocrinal effects of bone on energy metabolism (18). Moreover, an unexpected result of investigations on osteocalcin, a non-collagenous protein that is produced by osteoblasts and is involved in mineralization and calcium homeostasis, showed that osteocalcin gene knockout (OC-/-) animal models had an abnormal amount of visceral fat, thus providing the first evidence of a feedback loop between the skeleton and energy metabolism (19). This has led some researchers to conduct investigations to determine whether bone should be considered a ductless endocrine organ in addition to its classic functions of providing a framework to support the body and calcium homeostasis.
NOVEL METABOLIC ROLES OF OSTEOCALCIN
In 2007, a revolutionary reciprocal relationship between bones of the skeleton and energy metabolism was proposed, whereby leptin influences osteoblast functions and bone remodeling and OC in turn influences energy metabolism (20). The fact that only a limited number of osteoblast-specific genes encode functional molecules made it easy to examine whether transgenic constructs or deletion of these genes resulted in alterations in energy metabolism. Among these genes, the Esp gene encoding protein tyrosine phosphatase (OST-PTP) was of particular interest. Previous studies showed that the Esp gene is up-regulated following differentiation and matrix formation of primary osteoblasts and down-regulated in mineralizing osteoblasts (21-23). Deletion of the Esp gene in mice (Esp-/-) resulted in a high neonatal death rate from severe hypoglycemia associated with hyperinsulinemia and enhanced insulin sensitivity mediated by increased adiponectin production from adipocytes (20). This finding fostered the new hypothesis that osteoblast-specific OST-PTP may play a critical role in regulating glucose homeostasis.
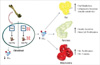
What makes the Esp gene and OST-PTP particularly interesting with respect to the relationship between bones and energy metabolism? In order to answer this question, a thorough understanding of the biophysical and biochemical properties of OC is required. OC is a vitamin K-dependent protein present in bones, and contains three glutamic acid residues that are posttranslationally modified by γ-glutamyl carboxylase (24, 25). This γ-carboxylation is essential for the protein to have a high affinity for mineral ions and enables the OC protein to attract calcium ions and incorporate these into hydroxyapatite crystals, which make up approximately 70% of bones (24). However, not all of the three glutamic acid residues in OC are fully carboxylated and incorporated into hydroxyapatite crystals, and the degree of carboxylation may vary. Uncarboxylated or undercarboxylated glutamic acid residues make OC susceptible to release from osteoblasts into the circulation (26). Hinoi et al. (27) demonstrated that leptin induces up-regulation of Esp expression on osteoblasts through sympathetic signaling without affecting overall production of OC, but causing a decrease in the serum concentration of uncarboxylated OC (OCuc). This study confirmed the report by Lee et al. (20) demonstrating increased concentrations of circulating OCuc in Esp-/- mouse models, besides providing further evidence that fat regulates bone metabolism. Although the exact mechanism is unclear, it is assumed that inhibiting OST-PTP expression negatively influences γ-car-boxylation of OC, thus promoting the release of OC into the circulation in the form of OCuc. Serum OCuc has been negatively associated with bone mineral density at measured bone sites (28-33). Surprisingly, serum OCuc appears to be sufficiently physiologically active to regulate energy metabolism, allowing bone to function as an endocrine organ (Fig. 2) (20, 34). Esp-/- mice representing OCuc-gain-of-function would be appropriate to determine the beneficial consequences of a high serum concentration of OCuc ([OCuc]) on energy metabolism.
Lee et al. (20) compared the effects of different [OCuc] on energy metabolism in genetically modified animal models (OC-/- and Esp-/-) and found that Esp-/- animals with significantly higher [OCuc] appeared to be protected from obesity, as evidenced by higher energy expenditure and reduced body weight and fat accumulation after being fed a high fat diet compared with wild-type (WT) and OC-/- models. Esp-/- models also exhibited lower serum levels of triglyceride and free fatty acids, suggesting that OCuc plays a role in lipid metabolism. OCuc also improved the insulin secretion capacity of the pancreas as measured by glucose-stimulated insulin secretion test and insulin tolerance test. This was evidenced by increased expression of MKI-67 and cyclin D, markers for cellular proliferation, in samples of pancreatic cells and a subsequent increase in pancreatic β-cell mass and area, and by increased secretion of adiponectin, an insulin-sensitizing adipocytokine, in Esp-/- animal models. OCuc also appeared to improve glucose tolerance determined by a glucose tolerance test and glucose infusion rate test. It is well known that reduction of mitochondrial content results in decreased mitochondrial function and leads to impaired energy metabolism and insulin resistance (35-37). Molecular and morphological analyses of the gastrocnemius muscle of Esp-/- animal models by Lee et al. (20) revealed an almost threefold increase in mitochondrial area and concurrent increase in the level of proteins associated with mitochondrial biogenesis including Mcad, PPAR-γ, acyl-CoA, UCP2, PGC1α, and NRF1 (38), suggesting that mitochondrial content and function were enhanced by OCuc. In contrast, OC-/- models displayed opposite characteristics of impaired lipid and glucose metabolism, and decreased insulin secretion and sensitivity (20). Thus, the revolutionary finding that OCuc positively regulates energy metabolism through increased fat metabolism, energy expenditure, insulin secretion capacity of pancreas, and release of adiponectin from adipocytes, and by inducing mitochondrial proliferation and function, established a complex crosstalk between bones and adipocytes in energy metabolism.
Evidence that OCuc enhances energy metabolism was confirmed not only in OC-/- mice exhibiting obesity, hyperglycemia, glucose intolerance, and insulin resistance, but also in WT mice (34). Ferron et al. (34) showed that OCuc had similar effects on energy metabolism in WT mice fed either a normal diet or a diet favoring obesity and type 2 diabetes after administration of OCuc. This study is especially notable as it proposes a pharmacological therapeutic potential of OCuc for obesity and type 2 diabetes. OCuc may also provide a novel treatment for sarcopenia, the loss of muscle mass and function with aging, when we consider the facts that activation of apoptosis initiated by mitochondrial dysfunction contributes to sarcopenia, and that OCuc induces mitochondrial proliferation and functional enhancement by increasing expression of proteins associated with mitochondrial biogenesis and functions (38, 39).
OSTEOCALCIN AS A POSSIBLE REGULATOR OF ENERGY METABOLISM IN HUMANS
Although an association between OC and energy metabolism has been demonstrated in mice, it is necessary to address the question of whether these metabolic functions of OC also exist in humans. Prior to the identification of the novel metabolic roles of OCuc in 2007, human studies performed over several decades reported markers of low bone turnover, including OC, in diabetic patients (40-42). Although OC has been extensively investigated in bone biology and related fields, it was not until very recently that OCuc was investigated in humans with respect to its metabolic functions. Im et al. (43) reported a significant reduction in [OC] among type 2 diabetic patients compared with normal glucose and impaired fasting glucose groups. These authors also showed significantly decreased fasting glucose and HbA1c levels in the highest quartile group for [OC] compared with the lowest quartile group in a study involving 339 Korean postmenopausal women (43). This inverse relationship was confirmed in studies on 1010 elderly Swedish men by Kindblom et al. (44) and studies on 380 elderly males and females by Pittas et al. (23). Human studies commonly reported an inverse relation between [OC] and fat mass and plasma glucose level; multivariate analyses by Im et al. (43) and Kindblom et al. (44) showed that [OC] is an independent negative predictor of plasma glucose, and Pittas provided the first demonstration that [OC] is inversely associated with the homeostasis model assessment for insulin resistance (HOMA-IR) in humans (23). Although [OCuc] was not measured, such observations suggest a potential metabolic role of OCuc in glycemia and adiposity in humans similar to that in the animal studies. It, however, would be premature to conclude a causal effect of OCuc on the regulation of energy metabolism in humans as the above studies investigated only correlations. Furthermore, these studies are limited by the fact that [OC] has been reported to vary with multiple factors including nutritional status, age, gender, smoking status, physical fitness levels, and season of the study (45, 46).
LIMITING FACTORS FOR SERUM OSTEOCALCIN LEVEL
As mentioned above, [OC] varies with multiple factors such as nutritional status, age, gender, smoking status, physical fitness levels, and season of the study (45, 46). Of these, vitamin K status is especially important because vitamin K is a cofactor for γ-glutamyl carboxylase responsible for γ-carboxylation of OC, thus vitamin K reduces the release of OC into the general circulation while increasing calcification of the bone (24, 25, 47). An inverse relationship between [OCuc] and vitamin K status has been reported in both human and animal studies (45-49), and consequently [OCuc] has frequently been used as a sensitive marker of vitamin K nutritional status (50). However, a recent study by Yoshida et al. (51), observed that high vitamin K1 (phylloquinone) intake is associated with increased insulin sensitivity and better glycemic status in the fed state. This observation further complicates the complex crosstalk between vitamin K, [OCuc], and glucose metabolism as it was expected that vitamin K would not positively influence glucose metabolism since it promotes γ-carboxylation of OC and thus decreases [OCuc]. Another vitamin to consider with regard to bone metabolism is vitamin D, which is known to affect osteoblasts. The biologically active and most widely used marker of vitamin D status is 1a,25-dihydroxyvitamin D3 (1a,25-(OH)2D3) (52). It is believed that 1a,25-(OH)2D3 positively influences differentiation and mineralization of osteoblasts via the vitamin D receptor on osteoblasts (53, 54). Although this relationship has not been further investigated, previous studies reported that 1a,25-(OH)2D3 increased overall OC production and showed an inverse correlation between [OCuc] and 1a,25-[OH]2D3 (46, 52-55), suggesting that 1a,25-(OH)2D3 has a bimodal effect on bone health and OCuc-mediated energy metabolism. Well-controlled studies on the crosstalk between vitamins, [OC], [OCuc], and energy metabolism are needed to better understand the relationship between bones and energy metabolism.
SUMMARY AND CONCLUSION
The reciprocal relationship between bones and energy metabolism that arose from the recent identification of novel metabolic roles of OCuc has excited integrative physiologists, especially those in osteology and endocrinology. This review mainly discusses this reciprocal relationship, or feedback loop, from a hormonal regulation perspective, and reviews newly discovered metabolic functions of osteocalcin. Leptin exerts antagonistic bimodal effects on bone remodeling through two distinct hypothalamic pathways that function via the sympathetic nervous system and CART, thereby acting directly on OC-secreting osteocytes, or osteoblasts. Conversely, bones participate in energy metabolism through hormonal regulation whereby OC within the bones is released into the circulation in the form of OCuc to increase energy expenditure, increase insulin secretion capacity by enhancing pancreatic β cell proliferation, increase insulin sensitivity by increasing adiponectin secretion, and reduce fat mass. This revolutionary discovery inspired Wolf (18) to propose that the large surface of the skeleton represents an excellent site of hormone synthesis. It is noteworthy that the osteoblast-specific phosphatase encoded by the Esp gene, OST-PTP, plays a critical role in this hormonal regulation. The serendipitous finding that mice lacking an osteoblast specific protein are abnormally obese has led to fundamental progress in related fields and addressed many research questions; however, many questions remain. First of all, the role of leptin in bone remodeling in humans is not yet clearly defined despite clarity in animal studies. The association between obesity and leptin resistance (56) explains the paradoxical observations that obese individuals generally have a lower prevalence of osteoporosis when compared with non-obese individuals and that leptin secretion is proportional to fat mass. However, inconsistent associations between serum leptin levels and bone mineral density from human studies have been reported (positive [57, 58]; negative [59-61]; or no association [62-64]) and the association between intracerebral leptin levels and bone mineral density in humans has not been investigated yet. Therefore, data obtained to date complicates interpretation of the effects of leptin on bone biology in humans and emphasizes the necessity for further investigations. Secondly, while positive influences of the skeleton on energy metabolism are causal in animal models, this remains unproven in humans although a positive correlation between OC and energy homeostasis was reported in very recent human studies. With respect to future research topics, Lee and Karsenty proposed identification of the OC receptor, the nature of OC regulation, and the interactions between this novel pathway and other pathways regulating energy metabolism (65). Other research topics include elucidating the crosstalk between nutritional status, OC, and energy metabolism, and investigating the contribution of OC in the development of treatments for components of metabolic syndrome such as obesity and both type I and II diabetes, and sarcopenia. Thorough and objective research on these topics is expected in related scientific fields over the following years.
 XML Download
XML Download