

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The addition of a methyl group to CpG dinucleotides is an essential contribution to an epigenetic mechanism by which the mammalian genome is programmed to establish different sets of genes expressed in different tissues (1, 2). The maintenance DNA methyltransferase has a replication error rate of up to 5% during somatic cell division (1). In contrast to the identical DNA sequence that is found in every cell, variation in DNA methylation can result in a tissue-specific pattern and is necessary to meet the demands of the later-life environment. Individuals that become infected with Helicobacter pylori undergo a series of discrete gastric mucosal changes during their lifetime (3). In a mouse model infected with H. pylori, long-standing gastric mucosal damage and the resultant mucosal changes are known to precede the development of gastric cancer (4). Therefore, in non-cancer individuals, repeated mucosal injuries are likely to induce adaptive methylation changes that enable efficient wound healing and act as a measure against cancer evolution.
The methylation-variable sites that locate in promoter or noncoding neutral regions have demonstrated gradual methylation changes associated with the aging or long-term adaptation process (5). A previous study proposed the presence of transitional-CpG sites between the unmethylated promoters and nearby densely methylated retroelements in order to describe the complexity of variable methylation in gene control regions (6, 7). The transitional-CpG sites at the margin of the CpG islands and at the non-island CpG sites around the transcription start sites have been found to be either under- or over-methylated in a tissue-specific manner as well as to be methylated to various degrees in the same tissue type (7). This suggests that the methylation-variable sites nearest to the transcription start sites may serve as epigenetic markers for adaptive DNA methylation.
There is a large body of evidence indicating that gene expression depends on communication between DNA methylation and histone modification that induces chromatin condensation (1). In addition, the dynamic chromatin structure is known to assemble transcription machinery on a master gene that enables the well-coordinated expression of tissue-specific genes and housekeeping genes (also known as the self-organization model) (8). In this regard, the methylation-variable site of a strongly expressed tissue-specific gene can influence the expression of the nearby gene as well as its related genes. Therefore, the methylation-variable sites of the key gastric-specific genes are expected to participate in the discrete mucosal adaptation and encapsulate the interactive methylation patterns that lead to mucosal changes, including gastric cancers.
In this study, the methylation profiles of 14 genes including the ulcer-healing genes, TFF1, TFF2, CDH1, and PPARG (9-11), in the normal gastric mucosa were delineated by examining the nearby methylation-variable sites. Both the TFF2 and PPARG genes were frequently undermethylated in the normal mucosa of ulcer patients. However, although the PPARG gene was undermethylated in the normal mucosa of cancer patients, the TFF2 gene was barely undermethylated, even when it was in close proximity to the cancer margin. In addition, a combination of over- or intermediate-methylated TFF2 and undermethylated PPARG genes was more common in stage-1 cancer patients than in healthy individuals (odds ratio [OR], 21.9) and non-invasive cancerous patients (OR, 8.9). These results suggest that the methylation-variable sites of the ulcer-healing genes may be accurate epigenetic markers for a high risk of gastric cancer.
MATERIALS AND METHODS
Normal gastric mucosa and pathologic tissues
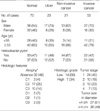
Seventy healthy individuals, 53 gastric cancer patients, 21 gastric non-invasive cancer patients, and 23 invasive gastric ulcer patients between September 2005 and July 2007 were enrolled in this study (Table 1). The control and patient groups all underwent either an endoscopic biopsy or a surgical resection at St. Paul's Hospital and St. Vincent's Hospital, The Catholic University of Korea. All subjects provided informed consent and the study was approved by the institutional review board (Document number 48, 'Genetic study of gastrointestinal cancer', January 28, 2005, St. Paul's Hospital, The Catholic University of Korea, Seoul, Korea). For the normal controls, two biopsy-tissue specimens were obtained from the antrum and the body of the normal stomach separated by a distance of 5 cm. The normal gastric mucosa of gastric ulcer, non-invasive cancer, and invasive cancer patients were endoscopically obtained 2 and 5 cm away from the lesion margin. Resected non-invasive and invasive cancer tissues were microscopically dissected in order to purify the normal epithelial cells within 1 cm of the lesion margin as well as the non-invasive and invasive cancerous cells.
To diagnose Helicobacter pylori infection, the paired biopsy samples collected 2-cm from the normal mucosal sites selected for methylation analysis were stained using the Warthin-Starry silver impregnation method. The grade of gastric atrophy was evaluated using the endoscopic atrophic border scale described by Kimura and Takemoto (12), which correlates with the results of the histological evaluations. Gastric non-invasive cancer and invasive cancer was diagnosed according to the Vienna classification of gastrointestinal epithelial neoplasia (13). Gastric ulcers that appeared malignant were defined based on an endoscopists impression of ulceration in the stomach and a microscopic examination showing no invasive cancer cells. The clinicopathological tumor stage was determined using the Tumor-Node-Metastasis (TNM) criteria (14).
Histological analysis of the normal mucosa confirmed that the biopsy tissues consisted primarily of normal gastric epithelial cells and showed no evidence of cancer cell invasion or significant inflammation. All of the microdissected cancer tissues contained a cancer cell content of ≥80%. Approximately 50 cells were digested in 1 µL of a Tween 20-Proteinase K lysis buffer after which a DNA isolation kit (A1120, Promega, Madison, WI, USA) was used to extract the genomic DNA according to the manufacturer's instructions.
Semiquantitative methylation analysis
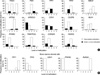
A search of the genes expressed in normal gastric mucosa and gastric cancers was made from eight SAGE (Serial Analysis of Gene Expression) libraries of four normal gastric mucosa and four gastric cancers that were obtained from a public database (http://cgap.nci.nih.gov/SAGE/). Highly expressed gastric genes (PGA5 and PGC) (15), ulcer-healing genes (TFF1, TFF2, CDH1, and PPARG) (9-11), cancer-associated genes (MLH1, CDKN2A, and RUNX3) (16, 17), weakly active genes (MSLN, ARRDC4, and DUPS6) (18-20), and non-gastric genes (KRT6A and TRAPPC2L) (21, 22) were selected to evaluate the methylation-variable sites. Based on a review of studies conducted to evaluate the transcription of each gene, the transcript number of individual genes counted in the SAGE data was found to accurately estimate a wide range of strong and weak transcription. The primer sites, sequences, and conditions are shown in Fig. 1, Table 2.
Each CpG amplicon site was analyzed by cloning and sequencing the common-polymerase chain reaction (PCR) products encompassing both the unmethylated and methylated CpGs, as previously reported (7, 16, 23, 24). Standard curves were then plotted for each methylation specific PCR (MSP) primer set using a mixture containing the methylated and unmethylated DNA clones at various ratios. The methylation status of the methylation-variable sites was scored at the following 5 levels by calculating the relative proportion of the methylation amplicon to the total intensity of unmethylation and methylation amplicons (7, 16, 23-25): 1 (0-20% methylation), 2 (21-40% methylation), 3 (41-60% methylation), 4 (61-80% methylation), and 5 (81-100% methylation) (Fig. 2). Because the number and position of methylated CpG dinucleotides were so heterogeneous in the methylation-variable sites, a wide range of methylation-variations was categorized as either under- and over-methylation based on an intermediate methylation level that was dominant in normal stomach samples (Fig. 3A).
The methylation-variable sites containing a low GC content and simple repeat sequence provided very limited sequences for the design of MSP primers. Therefore, a stringent PCR condition using a low concentration of dNTP was necessary to enable the specific amplification of simple sequences. The use of a dTTP-radioisotope was an essential step in generating the sharp electrophoresis bands required for semiquantitative MSP analysis. The reproducibility of semiquantitative MSP analysis was ensured using the following protocols: 1) 10 µL of a PCR mixture contained 1 µCi of α-32P dTTP (PerkinElmer, Boston, MA, USA), 62.5 µM dATP, dCTP, and dGTP, 25 µM dTTP, 1 pM of primers, 0.1% Tween 20 and 0.3 unit of Taq polymerase, 2) a fresh genomic DNA of 10 to 20 ng/µL was used for bisulfite modification, 3) each primer set covering 3-5 CpG sites was designed to generate a small amplicon of ≤150 bp, 4) bisulfite-modified DNA was amplified by subjecting it to 32 PCR cycles under hot-start PCR conditions, and 5) each MSP primer set was found to reproduce the same range of band intensity in more than 95% of duplicated experiments using the same biopsy tissue DNA (7, 16, 23, 24).
Comparison of radioisotope-labeling and non-radioisotope methods
Fourteen methylation-variable sites per each bisulfite-modified template DNA were amplified concurrently using the same PCR mixture and machine (Fig. 2). A total of 28 MSP amplicons was electrophoresed simultaneously on polyacrylamide gels. The dTTP-labeling protocol resulted in the balanced amplification of the unmethylated and methylated low-CpG DNA and allowed the semiquantitative measurement of methylation levels. However, when the non-radioisotope PCR protocol was used on the same samples subjected to the same PCR conditions, weak signals, noisy backgrounds, and smearing bands were often observed. The real-time PCR protocols, which have been used to measure methylation-dependent amplification of the CpG-rich island sites, also showed unstable and weak amplicon intensities in the methylation-variable sites (26).
Statistical analysis
A Fisher's exact test was used to determine if there were any significant differences in the frequency of under- or over-methylation in the mucosa of normal stomach versus either normal gastric mucosa or pathologic tissues obtained from ulcer, non-invasive cancer and cancer patients. The levels of methylation estimated in the mucosa of normal stomach and pathologic lesions were compared using a Mann-Whitney U test. The correlation coefficient was calculated to determine the concurrent methylation pattern between the ulcer-healing (CDH1, TFF1, and TFF2) genes and the other genes. The odds ratios and 95% confidence intervals (CI) were calculated using the relevant 2×2 contingency tables. Two-sided P values of <0.01 were considered to be significant.
RESULTS
Methylation-variations in the mucosa of normal stomach
The methylation status of the methylation-variable sites of 14 genes in the antrum and body of 70 normal stomachs was evaluated (Fig. 3A). Intermediate-level methylation was dominant for most of the genes examined in the normal stomach samples. Because the TFF1, TFF2, and CDH1 genes were methylated to various degrees in an age-dependent manner (Fig. 4A), the intermediate-level methylation of these three genes was relatively less frequent. When the normal stomach samples were divided into four age groups (≤44 yr [n=22], 45 to 54 yr [n=34], 55 to 64 yr [n=42], and ≥65 yr [n=42]), the TFF1 gene was more overmethylated in the older-aged group (≥55 yr) than in the younger-aged group (49% vs. 21%, respectively, P=0.002). The TFF2 gene was more undermethylated in the older-aged group (≥65 yr) than in the younger-aged groups (55% vs. 27%, respectively, P=0.002). The overmethylation of the CDH1 gene was most frequent in the age group that ranged from 45 to 54 yr (56% vs. 25%, respectively, P=0.012) and was associated with H. pylori infection (P<0.001, Fig. 4B).
Variations in methylation detected in normal stomach samples were analyzed to identify any possible relationships with the anatomic site, gender, age, and gastric atrophy (data not shown). The undermethylation of the TRAPPC2L non-gastric gene was more common in the body than in the antrum (36% vs. 4%, respectively, P<0.001) and more common in the older-aged group (≥55 yr) than in the younger-aged group (<55 yr) (29% vs. 4%, respectively, P=0.001).
Methylation-variations in the normal mucosa of ulcer, non-invasive cancer, and invasive cancer patients
The methylation status of the 14 selected genes was analyzed in normal mucosal tissues located 2 cm and 5 cm distal from 23 ulcerative, 21 non-invasive cancerous and 53 invasive cancerous lesions (Table 3). The methylation status of each gene was then compared between the paired normal mucosa (2 and 5 cm distances) of the pathologic lesions and the paired mucosa (antrum and body) of normal stomach samples. In stages 2, 3, and 4 cancers, 46 normal tissues were obtained from 29 cancer patients due to large-sized cancers. The normal mucosa of ulcer patients showed significant increases in the undermethylated PGC (20%, P=0.005), TFF2 (63%, P=0.001), MLH1 (13%, P=0.003), and PPARG (57%, P<0.001) genes. The overmethylated CDKN2A (43%, P<0.001) and RUNX3 (31%, P=0.003) genes were significantly common in the normal mucosa of non-invasive cancer patients. The normal mucosa of cancer patients showed significant increases in the overmethylated TFF2 (12%, P=0.004), KRT6A (29%, P<0.001), and CDKN2A (24%, P=0.003) genes as well as in the undermethylated PGC (16%, P=0.006), CDH1 (54%, P=0.005), DUSP6 (34%, P<0.001), MLH1 (12%, P=0.001), and PPARG (82%, P<0.001) genes.
Forty-eight normal tissues from stage-1 cancers and 46 normal tissues from advanced cancers were separately evaluated (Fig. 5A). Advanced cancer patients revealed that the TFF2 (15%, P=0.002), KRT6A (46%, P<0.001), and CDKN2A (26%, P=0.006) genes were significantly overmethylated in the normal mucosa, and that most of the other genes were undermethylated (P<0.01). In stage-1 cancer patients, only the PPARG gene was significantly undermethylated in normal gastric mucosa (P<0.001).
The methylation status of the PGA5, PGC, TFF2, PPARG, CDKN2A, and RUNX3 genes in the normal mucosa collected 2 cm and 5 cm distal from the three gastric lesions (ulcer, non-invasive cancer, and stage-1 cancer) were comparatively analyzed (Fig. 5B). The PGA5, CDKN2A, and RUNX3 genes tended to be increasingly undermethylated in the 2-cm mucosa of the three gastric lesions. The TFF2 gene was the most overmethylated or barely undermethylated in the two normal tissues of stage-1 cancer patients and the most undermethylated in the two normal tissues of ulcer patients.
Coordinate methylation patterns in normal gastric mucosa
The CDH1 gene with CpG island and the TFF1 and TFF2 genes without CpG islands in the normal mucosa of normal controls tended to be methylated concurrently with the methylation status of other CpG-island genes (ARRDC4, PPARG, and RUNX3) and non-island-CpG genes (PGC and MSLN), respectively (Fig. 6). In the normal mucosa of stage-1 cancer patients, only the methylation status of the TFF1 gene was positively associated with that of the PGC gene and the TFF2 gene was weakly associated with the PGC gene. In addition, in stage-1 cancer patients, the methylation of the TFF1 and TFF2 genes tended to be positively and negatively associated with that of the PPARG gene, respectively, although the relationships did not reach a statistical significance.
Odds ratios were estimated for the methylation patterns of the TFF2 and PPARG genes in the normal gastric mucosa of ulcer, non-invasive cancer, and stage-1 cancer patients (Table 4). The intermediate- or over-methylated TFF2 gene was more common in stage-1 cancer patients than in ulcer patients (85% vs. 37%, respectively; OR, 10.0; 95% CI, 3.7-27.2). The undermethylated PPARG gene was more common in cancer patients than in non-invasive cancer patients (80% vs. 36%, respectively; OR, 6.8; 95% CI, 2.7-17.5). A biopsy case showing the intermediate- or over-methylated TFF2 and undermethylated PPARG genes was defined as a discordant methylation pattern. When control individuals and stage-1 cancer patients were compared, the discordant TFF2-PPARG pattern was more common in stage-1 cancer patients (71% vs. 10%, respectively; OR, 21.9; 95% CI, 9.5-50.2) and the association with cancer patients was stronger in the older-age group (≥55 yr; 69% vs. 5%, respectively; OR, 43.6; 95% CI, 12.7-149.6). The discordant TFF2-PPARG pattern was more common in cancer patients than in ulcer patients (17%; OR, 11.5; 95% CI, 4.3-30.9) or non-invasive cancer patients (21%; OR, 8.9; 95% CI, 3.4-23.4).
Methylation-variations in non-invasive and invasive cancerous lesions
Of the six stomach-related or ulcer-healing genes examined in this study, only the TFF2 gene was significantly overmethylated in the cancerous lesions (15%, P=0.002) (Table 5). The PGC, TFF1, CDH1, DUSP6, and PPARG genes were largely undermethylated in both the non-invasive and invasive cancerous lesions (P<0.01). The MSLN and KRT6A genes were frequently overmethylated in both the non-invasive and invasive cancerous lesions (P<0.01). In the normal mucosa collected from within 1-cm of the non-invasive and invasive cancer margins, the methylation status of the genes showed a trend similar to that observed in the non-invasive and invasive cancerous lesions. However, the RUNX3 gene was significantly undermethylated in the 1-cm mucosa of non-invasive cancer patients, despite the common occurrence of overmethylation in non-invasive cancerous lesions and 2- and 5-cm normal mucosa.
DISCUSSION
In the normal gastric mucosa obtained from healthy individuals, the methylation-variable sites of ulcer-healing genes were concurrently methylated with other genes depending on the presence or absence of CpG islands (Fig. 6). These concurrent methylation patterns agree the results of other studies reporting that the epigenetic structure of tissue-specific and housekeeping genes are co-regulated to preserve a tissue environment (8). Both the TFF2 and PPARG genes were primarily undermethylated in normal mucosa of ulcer patients (Table 3). It is well known that the PPARG gene is involved in the gastric mucosal defense mechanism through up-regulation of TFF1 and TFF2 expression (9, 11). The concurrent methylation of the TFF2 and PPARG genes is compatible with wound healing and could mediate the mucosal defense system adequately in response to mucosal injuries. Therefore, the results of this study imply that the coordinate methylation of ulcer-healing genes is normally programmed in a gastric tissue environment to enable efficient recovery from mucosal damage.
The TFF1, CDH1, DUSP6, PPARG, and RUNX3 genes, which all play a role in stimulating mucosal growth or regulating the ulcer-healing process (9-11, 17, 20), tended to be extensively undermethylated in the normal mucosa of advanced-stage cancer patients (Fig. 5A). Meanwhile, the KRT6A and MSLN non-gastric genes without CpG islands in non-invasive and invasive cancer lesions as well as their 1-cm mucosa, which have a high cell growth rate, were found to be overmethylated. With respect to the methylation kinetics, previous studies evaluating the induction of cell differentiation have suggested that rapid cell division allows the undermethylation of the key tissue-specific genes to ensure cell proliferation and differentiation in a given tissue (23, 27). Consequently, the extensive undermethylation of the house-keeping and stomach-related genes may be a manifestation of the normal cell-intrinsic program that accelerates the mucosal growth in response to mucosal injuries.
The methylation profiles of the TFF1, TFF2, and CDH1 genes in the stomach of the younger-aged individuals (Fig. 4A) were different from those in the bone marrow samples (Fig. 3B). However, the three ulcer-healing genes in the stomach of older-aged individuals showed marrow-like methylation patterns, such as increases in overmethylated TFF1, undermethylated TFF2, and undermethylated CDH1 genes. Moreover, a previous study showed that H. pylori-infected gastric mucosa was associated with Polycomb proteins that regulate both DNA methylation and histone modification in pluripotent marrow cells (28). Therefore, it is likely that the gastric mucosa of older healthy individuals remodeled the epigenetic structure under the influence of H. pylori infection that promoted the recruitment of marrow-derived stem cells (4).
Because highly expressed TFF1 and TFF2 genes, which play a master role in mucosal repair (9), can self-organize a chromatin compartment for coordinate gene expression (8), it is likely that the methylation patterns of the two master genes influence their gene-specific chromatin structures as well as other gastric genes in the gastric mucosa. In this study, the TFF1 gene was tended to be methylated concurrent with the genes with and without CpG islands in normal stomach samples. The methylation status of the TFF2 gene was associated with that of the PGC and MSLN genes without CpG islands. In a recent study, the TFF1 gene was densely methylated in extraneous tissues and the TFF2, PGC, and MSLN genes were weakly methylated irrespective of their expression patterns (26). When applying the self-organization model to the stomach, the relative undermethylation of the TFF1 gene with dense-methylation site nucleates a transcription center and recruits a group of genes for the coordinated gene expression, and the TFF2, PGC, and MSLN genes with weak-methylation site subsequently participate in gene regulation. The weak methylation sites are likely to be variably methylated and selected to coordinate gastric-specific methylation patterns and gene expression in the gastric tissue environment.
The weak-methylation site of the TFF2 gene tended to be weakly or negatively associated with that of the PGC and PPARG genes in normal mucosa of stage-1 cancer patients (Fig. 6). The TFF2 gene was more methylated in stage-1 cancer patients than in ulcer patients, although the PPARG gene was significantly undermethylated in both cancer and ulcer patients (Table 4). This TFF2 methylation pattern indicates incomplete or aberrant coordination of highly expressed gastric gene, which may allow for the reactivation of marrow transcription patterns and cause the pluripotent stem cells to be vulnerable to neoplastic transformation. In this regard, the presence of discordant TFF2 methylation in the normal mucosa beyond the pathologic lesions is an indication of wide distribution of the cancer-prone progenitor cells (also known as field cancerization) (29).
A recent study suggests that the transcription-ready state of pluripotent marrow cells suppressing non-island-CpG genes is provided with the low-level methylation of CpG-island margins, while the gastric-specific genes lacking CpG islands in the stomach tissue are up-regulated with a concert effect of increasingly methylated CpG-islands (26). Additionally, gastric-gene expression was found to decrease in cancer tissues that reduced CpG-island methylation (Fig. 1, Table 5). Previous studies also have shown variable hypermethylation of CpG islands in H. pylori-infected mucosa of both non-cancer individuals and cancer patients, but a correlation between CpG-island methylation and gene silencing was obscure (30). Therefore, it is possible that the increased methylation of multiple CpG islands including the CDH1 gene up-regulates the gastric genes lacking CpG islands and leads to the adaptive differentiation of marrow stem cells.
Pluripotent marrow cells are known to express Polycomb proteins to promote DNA methylation based on the epigenetic memory module that drives multilineage differentiation (28). Therefore, the hypermethylated CpG islands appears to be a vestige of DNA methylation associated with the adaptive differentiation of marrow stem cells in extraneous tissues. The three cancer-associated genes with CpG islands (RUNX3, MLH1, and CDKN2A) were commonly overmethylated in the gastric cancer lesions (Table 5), but not in normal mucosa of stage-1 cancer patients. This is consistent with previous studies reporting that the hypermethylated genes common in cancer tissues are not common in the nearby normal gastric mucosa (30). The CpG-island methylation associated with the adaptation of marrow stem cells appears to involve the cancer-associated genes, which are not frequent to be detected in cancer-risk mucosa.
The methylation status of the RUNX3 gene that has a long CpG island varies greatly in the normal gastric mucosa dependent on the distance from the pathologic lesions. For example, the overmethylated RUNX3 gene was common in the 5-cm mucosa of non-invasive cancer and stage-1 cancer patients; however the undermethylated RUNX3 gene was common in the 1-cm mucosa of non-invasive cancer patients as well as in the normal mucosa of advanced-stage cancer patients (Fig. 5B, Table 5). This indicates that the tissue damage effect of gastric cancers could induce a subsequent undermethylation in overmethylated RUNX3 genes that preexisted in gastric cancer patients, which resulted in there being no significant difference in RUNX3 methylation between healthy individuals and stage-1 gastric cancer patients. In the normal gastric mucosa of non-invasive cancer patients, the CDKN2A and RUNX3 CpG-islands were the most overmethylated, with no single gene being significantly undermethylated (Table 3). These findings suggest that the variable methylation of CpG islands requires interpretation in terms of gene coordination associated with mucosal injury and H. pylori infection.
Individuals with gastric atrophy and mucosal injuries develop various clinical courses that range from a benign ulcer to non-invasive cancer and cancer or both during their lifetime (3). The methylation patterns of the ulcer-healing genes examined in this study suggest that epigenetic adaptation occurs in the gastric tissue environment and facilitates mucosal repair and regeneration during the individual's lifetime. In addition, genomic DNA obtained from a small biopsy tissue of the normal gastric mucosa showed a discordant methylation pattern of the TFF2 and PPARG genes, which was strongly associated with the normal mucosa of stage-1 gastric cancer patients. Therefore, these methylation-variable sites could serve as a sensitive marker for predicting cancer-prone gastric mucosa prior to the development of gastric cancer.









 XML Download
XML Download