

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
In sporadic colorectal cancer, various genetic changes, including adenomatous polyposis coli (APC), mismatch repair (MMR), Wnt-mediated, BRAF-mediated, cell biology-associated, and p53 alterations are concurrently observed (1). These mutations are interconnected to generate diverse pathways of colorectal tumorigenesis.
Activation of the Wnt pathway through mutation of APC triggers familial adenomatous polyposis coli (FAP) syndrome, and somatic mutations of APC are frequently observed in the majority of sporadic colorectal cancers (2). Inactivating mutations of APC stabilize β-catenin, which is translocated into the nucleus, leading to transcriptional upregulation of oncogenes, such as c-myc and cyclin D1, and promotion of tumorigenesis. β-catenin, the key component of the canonical Wnt pathway, is regulated by a multiprotein complex consisting of APC, Axin, and GSK-3β.
MMR defects occur due to gene inactivation and hMLH1 promoter methylation, frequently leading to microsatellite instability (MSI). RAF mutations may additionally contribute to colorectal carcinogenesis by upregulating anti-apoptotic properties through the RAS/RAF/ERK pathway (3). The RAF V600E hotspot mutation, occurring in approximately 10% of colorectal cancers, is strongly associated with the microsatellite instability phenotype (MSI+) (4). More specifically, RAF mutations are found specifically in MSI+ sporadic tumors that result from aberrant hMLH1 promoter methylation, but not germline mutations. An earlier investigation showed that APC or p53 mutations are more frequent in microsatellite stable (MSS) than MSI+ tumors, indicating a significant negative association between these alterations and microsatellite instability (5). On the other hand, nuclear β-catenin expression is more common in MLH1-expressing sporadic colorectal cancers, associated with tumor progression in these tumors (6).
Structural changes in extracellular matrix (ECM) proteins are a prerequisite for cell migration during tissue remodeling, accomplished by complex control of the expression and activities of matrix metalloproteinases (MMPs) (7). Overexpression of MMPs leads to degradation of ECM, an essential step for tumor invasion and metastasis (8). Particular groups of MMPs, i.e., gelatinases A and B, also known as 72 and 92 kDa type IV collagenase or MMP-2 and MMP-9 respectively, are of particular interest with respect to their roles in the development and progression of colorectal cancer (9). The metastatic capacity of colorectal cancer cells is closely associated with enhanced MMP production by the malignant cell type. Alternatively, angiogenesis controlled by angiogenic factors, such as the vascular endothelial growth factor (VEGF) family of cytokines, is critical in tumor growth and metastasis (10). VEGF-A and VEGF-B participate in tumor development during adenoma formation, while VEGF-C functions in the advanced stages of colorectal cancer (11).
ARK5, a member of the AMPK family, is activated by glucose starvation through Akt, and acts as a major factor in Akt-dependent cancer cell survival and migration via stimulation of MT1-MMPs in vitro (12). Carcinoembryonic antigen (CEA) participates in cellular differentiation and tumor invasion (13). S100A4, also known as metastasin or calvasculin, is overexpressed in primary colorectal cancers, compared with normal mucosa, and liver metastases (10).
In this study, we determined the association of known genetic changes of tumorigenesis in sporadic colorectal cancers with tumor growth and invasion. We additionally analyze the mRNA expression of invasion-associated genes, specifically, MMP2, MMP9, VEGFA, ARK5, CEA, and S100A4, in 92 tissue samples and 12 cell lines.
MATERIALS AND METHODS
Tumor samples and DNA/RNA preparation
Fresh colorectal tissues and lymphocytes from 92 patients were obtained between 2006 and 2007 at the Asan Medical Center (Seoul, Korea). Of the 92 colon cancers, 32 were located in the right colon, 30 in the left colon, and 30 in the rectum. Tumor tissues and samples of the corresponding normal mucosa at least 5 cm from the tumor border were cryopreserved in liquid nitrogen immediately after acquisition. All neoplastic tissues were confirmed by histologic identification, and peripheral blood lymphocytes from patients were obtained for genetic and molecular analyses. All patient samples were acquired with informed consent, and the study was conducted with the approval of the Institutional Review Board for Human Genetic and Genomic Research, in accordance with the declaration of Helsinki. Twelve human colorectal carcinoma cell lines, specifically, RKO and SW48, HT29, SW480, and HCT116 from the American Type Culture Collection (Rockville, MD, USA), KM12c, WiDr, DLD1, HCT15, Caco2, and LoVo from the Korean Cell Line Bank (Seoul, Korea), and AMC5 established in our laboratory, were maintained. HeLa and CCD841 control cell lines were provided by Yonsei University (Seoul, Korea). Cell lines were maintained in their respective media supplemented with 10% fetal bovine serum and antibiotics, according to standard protocols. DNA was extracted using the standard method and the corresponding cDNA synthesized by amplification with random primers and SuperScript II RT (Invitrogen, Grand Island, NY, USA).
Genetic alterations associated with colorectal tumorigenesis
An automated DHPLC instrument (WAVE™, Transgenomic, Omaha, NE, USA) was employed to examine mutations in APC exons 1-14 of genomic DNA from tumor tissues and cell lines. We used the protein truncation test (PTT) to identify the translation-terminating mutation in exon 15 (14). 'Hot spots' or possible pathogenic mutation sites were determined for other genes, specifically, RAS codons 12 and 13 and RAF codon 600 (17). MSI status was evaluated on the basis of the Bethesda panel (BAT25, BAT26, D5S346, D2S123, and D17S250). To assay for hMLH1 methylation, we analyzed the 3' small region close to the transcription start site, which is invariably associated with absence of hMLH1 expression in colorectal tumors, in addition to the 5' promoter region (15).
Immunohistochemistry and immunofluorescence
Paraffin-embedded tissue cores of carcinomas were used for tissue microarray construction, and arrayed onto recipient paraffin blocks with a precision instrument (Beecher Instrument, Sun Prairie, WI, USA). Tissue array blocks containing core cylinders were subjected to immune staining based on the labeled streptavidin-biotin method using the DAKO LSAB® kit (DAKO, Carpinteria, CA, USA). Protein markers included Wnt-activated alterations (β-catenin, Axin2, GSK3β), mismatch repair proteins (MLH1, MSH2), MEK, CEA, and p53 (antibody details available on request). For all samples displaying negative immune staining on a tissue microarray, whole tissue sections were repeatedly assayed. Negative staining was mainly classified into two groups, specifically, no to weak staining (≤10%) and no staining. The former group included GSK3β, MLH1 and MSH2 (nuclear), and p53, and the latter contained Axin2 and MEK. For β-catenin and CEA, cytoplasmic and no or apicoluminal staining were interpreted as negative, whereas nuclear and diffuse cytoplasmic staining were defined as positive, respectively. For immunofluorescence, cells cultured on 8-well chamber slides (Lab-Tek, Nunc, Rochester, NY, USA) were treated with primary antibodies, followed by FITC-conjugated secondary antibodies (goat anti-mouse, Sigma, St. Louis, MO, USA; goat anti-rabbit, Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA). Images were collected using a laser scanning confocal microscope (Leica Microsystems GmbH, Wetzlar, Germany).
Real-time reverse transcription-polymerase chain reaction (RT-PCR)
ARK5, CEA, MMP-2, MMP-9, S100A4, and VEGFA gene copy numbers were analyzed by RT-PCR (primer sequences available on request). Quantitative real-time RT-PCR was performed on a LightCycler (Roche, Mannheim, Germany) using the Fast Start kit, according to the manufacturer's instructions. Upregulation was determined when differential expression of mRNA (tumor/normal epithelium) was over one-fold.
Gelatin zymography assay
MMP-2 and MMP-9 activities in culture media were examined on a gelatin zymography. Concentrated conditioned media (×10) mixed with sample buffer were electrophoresed on a 10% sodium dodecyl sulfate-polyacrylamide gel with 0.1% gelatin (Invitrogen, Grand Island, NY, USA) incorporated as a substrate for gelatinolytic proteases under non-reducing conditions at 125 V for 2 hr. The gel was incubated at 37℃ for 16 hr in fresh developing buffer, and stained with 0.5% Coomassie brilliant blue R-250 (Bio-Rad, Hercules, CA, USA). Bands on the gels were measured using densitometry analyses (Bio-Rad).
Invasion and migration assay
Twenty-four well Transwells containing 8 µm pores coated with Matrigel (50 µL/well; BD Biosciences, Palo Alto, CA, USA) were used to assess cell invasion. Cells (5×105) in serum-free medium were seeded in duplicate in the upper chamber and 3T3 fibroblast-conditioned medium as a chemoattractant was placed in lower wells. Chambers were incubated at 37℃ in 5% CO2 for 24 hr. Invading cells on the lower surface of the filter were fixed with ice-cold methanol, and stained using 0.5% crystal violet. For the migration assay, cells were incubated in chambers without Matrigel to examine migration potentials using a similar procedure as the invasion assay.
Statistical analysis
Genetic and molecular alterations as well as invasion-associated gene expression were compared with each other or various clinicopathologic parameters by cross-table analysis using Fisher's exact or Pearson's chi-square-tests, depending on statistical validity. The correlations between differential mRNA expressions of invasion-associated genes were assessed using the significance of the product-moment correlation test. Numerical variables or differential gene expression of independent groups were examined using the unpaired Student's t-test or analysis of variance (ANOVA) with least-squares deviation (LSD) verification. The significance level was adjusted at 5% for each analysis, and all calculations were performed using SPSS software (ver.13, SPSS Inc., Chicago, IL, USA).
RESULTS
Genetic changes in colorectal cancer tissues and cell lines
Alterations in p53 were the most frequent (71%), followed by Wnt-activated alterations (48%), and MMR defects (21%). APC or Wnt-activated alterations occurred in 66% (59/90 patients), whereas MMR defects or RAF-mediated alterations were identified in 47% (42/90 patients). The crossover rate between APC/Wnt-activated and MMR/RAF-mediated alterations was 26%. Colorectal tumorigenesis pathways involving distinct genetic and molecular changes were compared in terms of biological features (Table 1).
APC mutations did not differ among various tumor locations, while MMR defects were predominantly identified in right colon cancers (P=0.002). RAF V600E mutation was exclusively found right colon cancers (P=0.02), while RAS G12D mutation was significantly less frequent in rectal cancers (P=0.02). The crossover pathways between APC/Wnt-activated and MMR/RAF-mediated alterations were significantly more common in right or left colon cancers than rectal cancers (P=0.018). In terms of biological characteristics, MMR defects were three times more frequent in poorly differentiated or mucinous adenocarcinomas compared to well or moderately differentiated adenocarcinomas (P=0.027). Wnt-activated and p53 alterations were identified in all 12 colorectal cancer cell lines by immunofluorescence analysis. Crossovers between APC/Wnt-activated and MMR/RAF-mediated alterations were observed in nine of the cell lines, i.e., DLD1, HCT15, HCT116, HT29, KM12c, LoVo, RKO, SW48, and WiDr.
mRNA expressions of invasion-associated genes in colorectal cancer tissues and cell lines
Differential mRNA expressions (tumor/normal epithelium) of four invasion-associated genes, i.e., ARK5, MMP2, MMP9, and S100A4 were closely associated among them. MMP2 was closely related to those of MMP9 (P<0.001). ARK5 expressions were closely associated with MMP2, MMP9, and S100A4 expressions, whereas S100A4 expressions were additionally associated with VEGFA. Otherwise, CEA expressions did not correlated with other invasion-associated genes. Enhanced differential mRNA expressions of several invasion-associated genes were related to tumor progression and invasion (Table 2). Enhanced MMP2 expressions were more frequently identified in colon cancers than rectal cancers (P=0.046). Advanced cancers were associated with enhanced CEA expressions and poorly differentiated and mucinous cancers with enhanced VEGFA expressions (P=0.034 and 0.042, respectively).
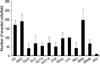
Enhanced mRNA expressions of most invasion-associated genes were relatively evident in AMC5 (ARK5, MMP2, MMP9, and VEGFA) and SW48 (ARK5, CEA, MMP2, MMP9, and VEGFA) cell lines (Fig. 1). KM12c and SW480 cell lines displayed increased S100A4 mRNA expression, whereas WiDr cell line did VEGFA mRNA overexpression.
mRNA expressions of invasion-associated genes in relation to genetic alterations
In sporadic colorectal cancers, differential mRNA expressions of invasion-associated genes were examined to correlate with genetic alterations (Table 3). Enhanced expression of ARK5 was more frequent in tumors with RAF-mediated alterations or crossover pathways carrying APC/Wnt-activated and MMR/RAF-mediated alterations than those without them (P=0.001 and 0.003, respectively). MMP9 expression tended to be increased in tumors with no p53 alterations (P=0.052). However, no significant association was identified in the limited number of colorectal cancer cell lines.
Invasion and migration assay in colorectal cancer cell lines
Activities of MMP-2 and MMP-9 were assayed using quantitative zymography (Fig. 2). Both active MMP-2 and active MMP-9 (62 and 82 kDa, respectively) were identified in all colorectal cancer cell lines except Caco2 and WiDr. Invasion of tumor cell lines was examined using a modified Boyden chamber (Transwell coated with Matrigel) (Fig. 3). Invasion of tumor cells was significantly greater in AMC5 and SW48 cell lines, compared to Caco2, RKO and WiDr (P=0.001-0.004). We additionally assessed in vitro migration using the Transwell migration assay. While all cell lines generally exhibited remarkably reduced migration compared with control 3T3, those displaying significant invasion generally maintained migratory activity as well (data not shown).
DISCUSSION
Previous investigations have disclosed various molecular changes in sporadic colorectal cancers, with APC, MMR, Wnt-activated, RAF-mediated, cell biology-associated, and p53 alterations accounting for approximately 80% of all occurrences (1). In our study, APC/Wnt-activated and MMR/RAF-mediated alterations constituted the major pathways of colorectal tumorigenesis (observed in 66% and 47% of patients, respectively). Alterations in p53 occurred in more than two-thirds of the patients in our study. Overexpression of mutant p53 has been reported in 5% to 32% of colorectal adenomas and 45% to 67% of carcinomas (16). In addition, Wnt-activated and p53 alterations were identified in all 12 colorectal cancer cell lines.
In our experiments, MMR defects were closely associated with right colon, poorly differentiated and mucinous cancers. Several investigators have reported correlations of right colon and less differentiated tumors with MMR defects in sporadic and hereditary colorectal cancer (17, 18). In our study, RAF mutation was exclusively identified in right colon cancer, while RAS mutation was significantly less in rectal cancer compared with colon cancer, possibly explaining unidentified differences of tumorigenesis between left colon and rectal cancers. Contrarily to our finding, one recent study demonstrated left colon cancer as a significant factor for the absence of RAS-signaling alteration (19). Our precedent study recognized rectal cancer as an attenuated phenotype of left colon cancer, with similarities in MMR defects, RAF-mediated and p53 alterations (20). Additionally, complex tumorigenesis accompanying the crossover between APC/Wnt and MMR/RAF alterations was more frequent for colon cancers than rectal cancers. In this particular context, MMR defects may trigger tumorigenesis by targeting transcriptional regulation genes, including Wnt and RAF signaling pathways.
We examined the mRNA expression of six invasion-associated genes: MMP2, MMP9, VEGF, ARK5, S100A4, and CEA. Most of these genes exhibited correlated expression among them, indicating synergistic interactions in vivo. Overexpression of MMPs leads to the degradation of ECM, an essential step for tumor invasion and metastasis (8). In human colorectal adenocarcinomas patients, MMP9 mRNA and plasma protein levels also increased, compared with normal individuals (21). Our results disclose a close correlation of MMP2 and MMP9 mRNA expressions in colon cancers. Previous investigations demonstrate that MMP-2 and MMP-9 increase in colorectal cancers with infiltrative growth, poor differentiation, and liver metastasis (22, 23). Our finding, shown significant upregulation of MMP2 mRNA in colon cancers compared with rectal cancers, appears to be further verified in terms of associated molecular changes using accumulative cases. Until now, several studies have not presented any difference of MMP2 activity between colon and rectal cancers (24).
Cancer cell-derived VEGF exerts pleiotropic effects on preexisting endothelial cells and tumor neovascularization (25). During this angiogenesis switch, VEGF regulates endothelial cell proliferation, migration, invasion, survival, and branching morphogenesis. VEGF-A and VEGF-B function early in tumor development at the stage of adenoma formation, while VEGF-C plays a role in advanced disease, such as metastatic spread of colorectal cancers. In our study, upregulation of VEGFA transcription was correlated with poorly differentiated or mucinous colorectal cancers. Similarly, expression of the major VEGF receptor, pVEGFR2/KDR, in cancer cells was associated with large tumors and poor differentiation (25). Hypoxia commonly develops within solid tumors as one of the most potent stimuli of VEGF expression (26).
AMP-activated protein kinases (AMPKs) are a class of serine/threonine protein kinases activated by an increase in the intracellular AMP concentration. ARK5, an AMPK catalytic subunit family member, plays diverse roles in human malignancy, i.e., tumorigenesis, proliferation, survival, invasion, and metastasis (27). Overexpression of ARK5 results in MMP-2 and MMP-9 activation through MT1-MMP overproduction in some cancer cell lines (27). This finding may explain the concurrent differential mRNA expressions of MMP2 and MMP9 in our study, suggesting a relationship between invasiveness and coexpression of ARK5 and MMPs. In our study, ARK5 mRNA expression was associated with RAF-mediated and crossover pathways, possibly signifying invasive alterations. Recent reports show that RAF is a novel indicator of progression and aggressiveness in papillary thyroid cancer (28). While glucose transport dependent on AMPK is mediated via ERK, the molecular function of ARK5 in the RAF pathway remains to be verified. S100A4, a direct β-catenin/TCF target, induces migration and invasion in vitro and metastasis in vivo, and is thereby applicable as a prognostic indicator in metastatic colorectal cancers (29). However, we did not observe any association of S100A4 upregulation with tumor stage, histologic differentiation, and lymphovascular invasion. CEA is a known metastatic potentiator in colorectal cancer, which facilitates adhesion and metastasis (13). In our experiments, advanced tumor was associated with upregulation of CEA, consistent with previous finding (30).
Among the 12 colorectal cancer cell lines examined, AMC5 and SW48 displayed overexpression of invasion-associated genes most frequently, specifically, ARK5, MMP2, MMP9, and VEGFA in AMC5, and CEA in addition to these genes in SW48. Additionally, the two cell lines exhibited active MMP-2 and MMP-9 expressions, known to be exclusively found in metastatic tissues (22, 23). These cell lines consequently exhibited the greatest invasion and migration activities on a Boyden chamber assay. On the other hand, KM12c and SW480 showed S100A4 upregulation with moderate invasion and migration. The biological aggressiveness data obtained support a strong correlation between the invasive behavior of tumor cells in vitro and in vivo, thus validating this assay as an objective measure of invasive potential (8). The six invasion-associated genes in this study may therefore be employed as a set of predictive markers for screening aggressive colorectal cancers, following verification in the clinical setting.
Conclusively, sporadic colorectal cancers undergo diverse molecular and genetic changes during tumorigenesis, including canonical and crossover pathways, namely, APC-Wnt alterations, MMR-RAF alterations, and crossover between the two. Specific pathways are closely associated with characteristic tumor growth and invasion, for example, RAF-mediated and crossover alterations with colon cancers and ARK5A overexpression related to other invasion-associated molecules, possibly employed as a panel of predictive markers.





 XML Download
XML Download