

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Cancer cells are self-altering cell populations that often do not derive an appropriate response from the immune system. Lack of tumor antigen presentation may lead to a deficit in the capability of the host immune system to destroy immunogenic tumor cells (1). Dendritic cells (DCs) are the most potent antigenpresenting cells (APCs) that play major roles in innate and acquired immunity. Tumor antigen-loaded DCs have been successfully applied as therapeutic vaccines to induce specific immunity against cancer. Mature DCs are better at inducing anti-cancer immune responses in vaccinated patients than immature cells.
Various techniques have been developed for improving the yield of mature DCs and enhancing their immune responses against cancer. Among these, a streptococcal preparation, OK432 (Picibanil®), increases the efficiency of DC maturation and induction of antigen-specific cytotoxic T cells in colorectal cancer cell lines (2). Interestingly, 'rapidly generated' fully mature DCs stimulated by a combination of OK432, low-dose prostanoids, and interferon-α (OPA) act as more potent immune enhancers than those stimulated via other methods (3). Here, we investigate the efficacy of early matured DCs treated with OPA as a fusion vaccine for the treatment of hormone-refractory prostate cancer.
MATERIALS AND METHODS
Generation of DCs from peripheral blood monocytes
Peripheral blood was drawn from healthy volunteers with informed consent, after the approval of the institutional review board (2007-12-159). Briefly, peripheral blood monocytes (PBMCs) were isolated from the buffy coat layer of blood by Lymphoprep® density-gradient separation (Axis-shield PoC AS, Oslo, Norway) after centrifugation at 1,800 r.p.m. Next, cells were incubated in culture flasks at 37℃ for 2-3 hr in AIM-V (GIBCO) medium. Aliquots of harvested cells were cryopreserved in 90% heat-inactivated fetal bovine serum (FBS) (GIBCO, Carlsbad, CA, USA) plus 10% dimethylsulfoxide (DMSO) (Sigma-Aldrich Corp. St. Louis, MO, USA). The remaining adherent cells were cultured in AIM-V medium supplemented with interleukin-4 (IL-4) (40 ng/mL) and GM-CSF (50 ng/mL) (R&D Systems, Inc., Minneapolis, MN, USA). Fresh medium was added up to half of the volume daily. Three sets of DC stimulatory agents were prepared, specifically, TNF-α only (20 mg/mL) (T only set), TNF-α (20 mg/mL) plus lipopolysaccharides (LPS) (1 µg/mL) (TL set), and OK432 (0.1 KE/mL) (Chugai Pharm. Co. Ltd., Taipei, Taiwan), prostaglandin E2 (50 ng/mL), and interferon-α (500 IU/mL) (OPA set). For conventional maturation of DCs, stimulatory agents were added on day 7 of culture. For early maturation, the three sets of agents were added on day 3 of culture. Using the six maturation methods combined the three sets of stimulating agents with two time-points (early and conventional), the efficacies of the mature dendritic cells were compared (Fig. 1).
Phenotype analysis and yield of mature DCs
Cells were incubated with FITC-conjugated antibodies against MHC class II (HLA-DR), costimulatory molecules (CD80 and CD86), and DC maturation markers (CD83, CCR7). PKH26, a prostate cancer marker, in a red fluorescent cell linker kit, was used to validate cancer cells. Using these markers, cell phenotypes were analyzed with FACS (Fluorescent activated cell sorter-FACSCalibur™; BD Immunocytometry Systems, San Hose, CA). The yield of mature DCs obtained with each of the six methods was compared via FACS analysis detecting CD86, a marker for mature dendritic cells.
Tumor cell lines
A hormone refractory human prostate cancer cell line, DU145, was purchased from the American Type Culture Collection (ATCC, Manassas, VA). The cell line was maintained in RPMI1640 medium, and used for fusion vaccines and cytotoxicity assays.
DC/tumor hybridization
DU145 cells were subjected to heat shock (42℃) for 2 hr, followed by recovery for 24 hr at 37℃ (4). After the pretreatment, cells were mixed with mature DCs prepared using the six maturation methods in a 50 mL tube at a ratio of 1:2. For hybridization, polyethylene glycol (PEG) (PEG1500, Roche Diagnostics, Mannheim, Germany) was pre-warmed to 37℃, and added directly to the mixed cell pellet. Following hybridization, 25 mL of 37℃ AIM-V medium was added slowly to cells. The DC-DU145 hybrids were characterized by FACS with the red fluorescent cell linker kit, PKH26. DCs were characterized by directly labeled CD86-FITC (Serotec, Oxford, UK). For FACS analysis, DC-DU145 hybrids were washed with PBS, and incubated with antibodies for 30 min at room temperature. Each fusion ratio of the hybrids in the six combination sets was analyzed and compared.
Generation of tumor-specific T cells
T cells were thawed and treated with IL-7 (20 ng/mL) and IL-12 (100 pg/mL) (R&D Systems Inc.). After overnight incubation, naïve T cells (responder cell) were stimulated with hybridized and non-hybridized DCs (stimulator cell) at the ratio of 10:1, respectively. Responder cells were cultured in X-vivo 15 medium supplemented with IL-7 (20 ng/mL), IL-12 (100 pg/mL) and 2% autologous plasma for 3 days. On day 4 of co-culture, IL-2 (20 IU/mL) (R&D Systems, Inc.) was added, and cytotoxicity against tumor cells analyzed.
T cell proliferation assay
Primary allogeneic mixed leukocyte reaction (MLR) was performed with hybridized and non-hybridized cells. Cells were cocultured with 105 allogeneic T cells/well in 96-well plates for 3 days. BrdU (1 µCi) was added to each well at 15-20 hr before harvest. Incorporation of BrdU (Roche, Penzberg, Germany) was measured colorimetrically.
Cytokine production assays
Supernatant fractions from PEG-treated and untreated DCs and DU145 tumor cell cultures were collected at 24-48 hr after hybridization, and analyzed for IL-12 secretion by DC-DU145 hybrids using ELISA (Mabtech AB, Stockholm, Sweden). ELISA was performed according to the manufacturer's instructions, and quantitated using a VERSAmax microplate reader (Molecular Devices Corp., Sunnyvale, CA, USA). Interferon-gamma (INF-γ) secreted by tumor-specific CD8+ CTL cells were also analyzed using ELISA (Mabtech AB).
Cytotoxicity assays
T cells stimulated with DC-DU145 hybrids were co-cultured with DU145 in 96-well plates for 24 hr at 37℃. The CytoTox-ONE assay® (Promega, Medison, WI, USA) provided a measurement of lactate dehydrogenase (LDH) released from cells with damaged membrane. All steps were performed according to the manufacturer's instructions. Briefly, after washing cells with assay medium (AIM-V), plates were centrifuged, and supernatant fractions were transferred to a flat-bottomed ELISA plate. The LDH detection mixture was added to each well and incubated in the dark for 10 min at room temperature. Stop solution was added, and the fluorescent signal (560EX/590EM) measured with a Fluorescence plate reader.
RESULTS
Phenotypes and yields of mature DCs
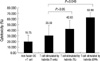
The mean number of harvested DCs was 8×106 cells per 50 mL of PBMC. HLA-DR, CD80, CD83, and CD86 were expressed at higher levels in mature DCs than immature cells. The DU145 cell line did not contain these molecules, and expressed PKH26 (data not shown). The yield of mature DCs was high at the early maturation time-point, irrespective of the type of stimulator (P=0.012). Moreover, yields tended to increase in the order of T only, TL, and OPA, in both early and conventional maturation settings. Specifically, the mean yield of mature DCs was 43.1% with the T only set, 47.8% with the TL set, and 58.7% with the OPA set in the early maturation setting, and 17.2%, 21.9%, and 27.4%, respectively, in the conventional setting (Table 1). Data from FACS analyses revealed a right shift in the figure in the same order, confirming this finding. FACS data for CD86 are presented in Fig. 2. Overall, the highest yield of early mature DCs was obtained with OPA.
Fusion of DCs and DU145
Fusion was performed with 2×106 DCs and 1×106 DU145 cells (2:1) stimulated with PEG solution. Hybridized cells were identified via dual positivity for PKH26-PI and CD86-FITC in FACS analysis (Fig. 3). In a conventional maturation setting, fusion ratios were comparable among cells stimulated with different types of maturation agents. However, under early maturation conditions, the fusion ratio of DU145 and DCs matured with the OPA set was the highest (28.7-60.8%), while that of DU145 and DCs matured with the T only set was the lowest (10.7-17.7%). The mean fusion ratios, calculated from the fraction of dual-positive cells, were 13.3%, 18.6%, and 39.9%, respectively (P=0.051) in the T only, TL, and OPA-treated groups. However, in the conventional maturation setting, fusion ratios were not different among the groups (9.9%, 11.2% and 13.1%, respectively). Cells generated at the early maturation time-point (3 days) additionally displayed superior efficacy to those obtained at the conventional time (7 days) in terms of fusion ratio (P=0.01) (Fig. 4).
Function of DC-DU145 hybrids
In view of the superiority of early maturation, the conventional maturation method was excluded in subsequent functional studies. The functions of hybrids were evaluated by measuring the level of IL-12, a cytokine secreted by mature DCs that is the most powerful known stimulator for inducing innate T cells to CD8+ T cells. Among the stimulatory agents, the OPA set was the most effective in promoting IL-12 secretion from the hybrids. Compared to the T only set, the OPA set was significantly more efficacious (P=0.021) but the TL set was not (P=0.083) (Fig. 5).
Enhanced T cell activation by DC-DU145 hybrids
DC-DU145 hybrids enhanced the proliferation of CD8+ T cells by sensitizing them to tumor antigens upon co-culture. Tumor-sensitized CD8+ cytotoxic T cells (CTL) produced a cytokine, IFN-γ. Similar to the IL-12 secretion function of DC-DU145 hybrids, T cell proliferation by OPA-treated DC-DU145 hybrids and IFN-γ secretion by sensitized CTLs were significantly superior to data obtained with hybrids of DCs matured using other methods (Figs. 6, 7).
Cytotoxicity of CTLs with different stimulations
In general, the cytotoxicity results were similar to those obtained from previous functional analyses. The cytotoxicity of CTLs sensitized with hybrids containing OPA-treated DCs was stronger than that of CTLs sensitized with hybrids containing T only-treated or TL-treated DCs (Fig. 8). The mean percentage of tumor lyses was 29.6% in the T only set, 42.8% in the TL set, and 62.9% in the OPA set. Compared to the T only set, the OPA set was significantly more efficacious (P=0.049) but the TL set was not.
DISCUSSION
Anti-cancer vaccines are a suggested treatment modality for advanced prostate cancer, and have been investigated in several clinical trials. The generation of high numbers of mature DCs is essential to obtain a potent anti-cancer vaccine, which is currently a challenging and unresolved issue (5). Various methodological improvement, including a range of DC maturation stimulators, maturation times, techniques of tumor antigen loading to DCs, and treatment of tumor cells before tumor antigen loading, have been explored to achieve this aim. While the evidence obtained to date is controversial (6, 7), the efficacy of the fusion procedure is superior to that of the tumor lysate-stimulating method (8, 9).
In the present study, we adopted recent findings on anti-tumor fusion vaccines for prostate cancer. Lundqvist and colleagues (10) suggested that an allogeneic tumor-dendritic cell fusion vaccine against prostate cancer is a potent stimulator of the broad prostate tumor-specific response. They employed TNF-α and IL-4 as maturation stimulating agents and conventional maturation times, and reported a hybrid yield of 10-25%. Our data confirm the superiority of the OPA combination as a DC maturation stimulator, and demonstrate that early maturation of DCs promotes the efficacy of antigen presentation to a greater extent than conventional 7-day maturation. Moreover, the overall yield of DC-DU145 hybrids obtained was 15-60%.
The feasibility of DC maturation as early as 2-day culture was reported by Tanaka and co-workers (11). They showed that 'rapid DCs' are equally as potent as '7-day conventional mature DCs' in inducing specific CTLs in vitro. Sakakibara and colleagues (3) reported similar results with colorectal cancer cell lines. Their data showed that quickly generated mature DCs were superior to those obtained with conventional 7-day maturation in terms of antigen-presenting ability. It was assumed that the shorter culture period gave rise to larger numbers of DCs, and quicker generation of DCs was beneficial for the maintenance of cell quality. The apoptosis rate of DCs increased from 10% to 25% over one week of culture (12). Our results support previous data showing that the yield of mature DCs is consistently and significantly higher in early cultures than the conventional 7-day time-period, irrespective of other conditions.
Upon culture with GM-CSF and IL-4, only immature DCs are harvested, which would induce immune tolerance in vivo (5). Maturation of DCs in vitro is inducible by inflammatory stimuli, such as TNF-α, IL-1β, LPS, unmethylated DNA containing CpG motifs, CD40 ligation, and contact sensitizers (13-16). Conventionally, TNF-α with or without LPS is employed for stimulating DC maturation. The LPS ligand combines with Toll-like receptor 4 (TLR4), a surface molecule of DCs, which, in turn, initiates intracellular signal transduction to the nucleus. Consequently, nuclear factor κB and Jun/Fos are activated at the nuclear level, and a series of gene expression steps mediate maturation of DC (17). Eventually, Th2 differentiation is induced by TNF-α and LPS. OK432, a lyophilized preparation of the Su strain of Streptococcus pyogenes inactivated with penicillin G, is a non-specific immunomodulator. The compound acts in combination with TLR4 to affect various immune functions, not only enhancement of cytotoxic cells, lymphocytes, macrophages, and NK cells, but also stimulation of cytokine secretion, such as interferons, TNF-α, and IL-2 (5). Several recent evidence show that OK432 is more effective than LPS in promoting the maturation of DCs (5, 18, 19). Our data also were similar to the results of previous studies. We estimated that the optimal concentration of LPS for maturation of DCs is 1 µg/mL. The maturation yield of DCs was decreased at a LPS dosage over 1 µg/mL, possibly resulting from cytotoxicity of LPS.
Conventionally, prior to hybridization, cancer cells are apoptosed via ionizing radiation ranging between 15 and 150 Gy to enhance fusion efficacy (10, 20). Instead of irradiation, we applied heat stress to tumor cells, which had a synergic effect on the induction of antigen-specific CTL with TLR-stimulated DCs, as suggested by Koido et al. (21). However, we have not compared the efficacy of heat-stress techniques with irradiation of DU145 cell lines as preparative procedures for hybridization in this analysis.
In most in vitro anti-cancer vaccine studies, the more efficacious DC maturation methods showed superior result in cytotoxicity against cancer cells, and functions of antigen-loaded DCs and sensitized CTLs. In view of the significant superiority of the early maturation procedure in the DC and DC-DU145 hybrid analyses, conventional maturation was excluded in subsequent functional studies. Among the DC maturation agents, OPA was the strongest immune modulator in cytokine secretion of the hybrids, T cell sensitization with cancer antigens, and T cell proliferation. While OPA induced the highest percentage of cytotoxicity, its superiority was not statistically significant in comparison to the other compound sets due to the small number of tests performed (only two samples). Additional studies are ongoing to acquire more data on cytotoxicity.
The main limitation of this study is that tumor specificity for DU145 in the cytotoxicity analysis was not clearly validated. To confirm this, the cytotoxicity of CTLs sensitized with DC-DU145 hybrids should be compared with that of the CTLs sensitized with the DC hybrids with cancer cell line originated from other organs. Another drawback was the significant diversity in cell quality of PBMCs among the donors. We abandoned data showing poor viability at the beginning of the study. Despite of this initial serum quality control, significant inter-personal variations were consistently observed throughout the analysis. To improve data quality, the initial amount of peripheral blood obtained from the donors may be increased from 50 mL to about 500 mL or higher, or PBMCs collected from volunteers injected with GM-CSF. Additional in vivo studies are required to apply the results to the clinical setting.
Enhancing the number and activity of the effecter cells using the presented methods in this study, the clinical results of cancer vaccine treatment for hormone refractory prostate cancer may be improved.
In conclusion, the antitumor effects of DC-DU145 hybrids generated with DCs subjected to early maturation with the OPA may be superior to the effects of those matured using conventional methods.








 XML Download
XML Download