

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Mesenchymal stem cells (MSCs) are capable of self-renewal and differentiation into lineages of mesenchymal tissues, including bone, cartilage, fat, tendon, muscle, and hematopoietic supporting stroma (1, 2). While the most abundant source of MSCs is the bone marrow (BM), MSCs can be obtained from other tissues such as peripheral blood, periosteum, muscle, adipose tissue, and the connective tissue of human adults (3).
MSCs from cord blood (CB) and the placenta may serve as alternative sources to adult MSCs. These potential alternative sources are important for a number of reasons, including the significant reduction in the number of MSCs with age and the high risk of viral contamination in the BM as well as the painful procedure required for the collection of BM MSCs. CB and placenta are potential alternative sources of MSCs; they are routinely discarded after delivery, and the ethical concerns and viral contamination issues are less of a problem. Several investigators have successfully isolated, expanded, and characterized the MSCs from CB and the placenta, and have evaluated their potential for differentiation into osteogenic, chondrogenic or adipogenic lineages (4-6).
In animal models, MSCs can be induced to circulate into peripheral blood under certain conditions, such as hypoxia (7). The chemotactic signals that guide MSCs to appropriate microenvironments or induce their circulation have yet to be identified. In hematopoietic stem cells (HSC), the predominant role of stromal derived factor-1 (SDF-1) and its receptor CXCR-4 is now well established (8). Some soluble factors have recently been reported to exert chemotactic effects on BM MSCs, including chemokines (9) and growth factors (10), however, their respective physiological relevance remains unclear. An improved knowledge of the chemotactic factors that affect MSCs would be of clinical interest since modulation of their activity could affect not only engraftment efficiency at damaged sites but also their mobilization into peripheral blood. Ponte et al. (11) showed that, in contrast to what is known about HSCs, a wide range of soluble factors exert significant chemotactic activity on MSCs and some growth factors are better chemoattractants than chemokines. Inflammatory cytokines such as tumor necrosis factor α (TNFα) can increase the sensitivity of MSCs to chemokines.
In this study, we characterized and compared the cytokine expression of BM-MSCs, CB-MSCs, and placental MSCs using a commercial human cytokine protein array assay to identify the protein expression profiles of the MSCs from BM, CB and the placenta. This study focused on the cytokines associated with the monocyte chemotactic protein-1 (MCP-1), soluble intercellular adhesion molecule-1 (sICAM-1) and SDF-1.
MATERIALS AND METHODS
Isolation of mesenchymal stem cells
MSCs from human BM, CB, and the amnion and decidua of the placenta were studied. The BM MSCs, in the first passage, were obtained from the Department of Orthopedic Surgery. The CB MSCs, in the first passage, were obtained from MEDIPOST Co. (Seoul, Korea). The human placentas (from clinically normal pregnancies, gestational age, 34-41 weeks) were obtained after vaginal deliveries or caesarean section births. All tissues were obtained with the approval of the Korea University Medical Center Institutional Review Board. The amnion and decidua were mechanically peeled from the placenta, and washed with phosphate-buffered saline (PBS) several times to remove excessive blood. The tissues were incubated with 1 mg/mL type I collagenase for three hours after being cut into small pieces (1 cm3) with scissors. The mononuclear cells in the collagenase were collected. After centrifugation, the cells were washed with PBS and resuspended in an α-minimum essential medium (MEM, Gibco BRL, Life Technologies, Merelbeke, Belgium) supplemented with 10% fetal bovine serum, and 10 ng/mL basic fibroblast growth factor (bFGF). The cells were seeded into a T75 flask. The cultures were maintained at 37℃ in a humidified atmosphere with 5% CO2. Three to five days after initiating the incubation, the small digested residues were removed and the culture was continued. The medium was replaced one to two times every week, every third to fourth day.
Flow cytometry analysis
The specific surface antigens of MSCs in the cultures of passage two to four were characterized by flow cytometry analysis. The cells in culture were trypsinized and stained with fluorescein isothiocyanate (FITC)- or phycoerythrin (PE)-conjugated antibodies against CD29, CD31, CD34, CD44, CD45, HLA-DR, CD73, CD90, and CD105 (Immunotech, Marseille, MN, U.S.A.). Thereafter, the cells were analyzed using a Becton Dickinson flow cytometer (Becton Dickinson, San Jose, CA, U.S.A.).
Adipogenic induction
The medium was replaced with adipocyte induction medium or control (stromal) medium, as previously described (12). The induction medium contained a low glucose Dulbecco's Modified Eagle's Medium (LG-DMEM) with 10% FBS, 200 µM indomethacin, 1 µg/mL insulin, 1 mM dexamethasone, 0.5 mM isobutylmethylxanthine (IBMX), 100 U/mL penicillin, 100 mg/mL streptomycin, and 0.25 mg/mL amphotericin B. After 3 days, the medium was changed to adipocyte maintenance medium that was identical to the induction media but without IBMX. The cells were maintained in culture for 14 days, with 90% of the maintenance media replaced every 3 days.
Osteogenic induction
The medium was replaced with osteoblast induction medium or control (stromal) medium, as previously described (12). Osteoblast induction medium contained DMEM (low glucose) with 10% fetal bovine serum, 10 mM β-glycerophosphate, 0.15 mM ascorbate-2-phosphate, 10 nM 1,25-(OH) 2 vitamin D3, 10 nM dexamethasone, 100 U/mL penicillin, 100 mg/mL streptomycin, and 0.25 mg/mL amphotericin B. The cells were maintained in culture for 21 to 28 days, with 90% of the media replaced every 3 days.
The assessment of cell viability and cell number in cultures
The cultured cells were detached from the culture dishes with 0.05% trypsin-EDTA (Gibco BRL, Life Technologies) at 72 hr of culture under different culture conditions. The cells were stained with trypan blue (Gibco BRL, Life Technologies), and the viable cells that did not stain were counted on a hemocytometer.
The focused protein array
The culture media were collected at different incubation periods. All focused protein array analyses were performed according to the manufacturer's instructions. Positive controls were located in the upper left-hand corner (two spots), lower left-hand corner (two spots) and lower right-hand corner (two spots) of each array kit. Each culture media was measured using the human cytokine array panel A (proteome profiler™) (R&D Systems, Minneapolis, MN, U.S.A.). Horseradish peroxidase substrate (Bio-Rad, Hercules, CA, U.S.A.) was used to detect protein expression and data were captured by exposure to Kodak BioMax Light film. Arrays were scanned into a computer and optical density measurements were obtained with the Image Pro Plus v 5.1 software (Media Cybernetics, Silver Spring, MD, U.S.A.).
Total RNA isolation and reverse-transcriptase reaction
RNA extraction and purification were performed using an RNeasy mini kit as described in the manufacturer's protocol (Qiagen, Valencia, CA, U.S.A.). The concentration of RNA was measured using a spectrophotometer (DU®530, Beckman, Fullerton, CA, U.S.A.), and the RNA quality was confirmed by agarose gels. A total RNA sample (2 µg/sample) was used with 20 µL of sample to generate cDNA using the SuperScript™ III First-Strand Synthesis System RT-PCR kit (Invitrogen, Milan, Italy). RNA was reverse-transcribed under the following conditions: 25 mM MgCl2, 10 mM dNTP mix, 10×RT buffer, 0.1 M DTT, 200 U of SuperScript™ III (Invitrogen), 40 U of RNaseOut, and 50 µM oligo d(T) primers in a final volume of 20 µL. The reaction was run at 65℃ for 5 min and at 50℃ for 50 min, and then the enzyme was heat inactivated at 85℃ for 5 min. For the real-time PCR reaction, 4 µL of reaction product were used.
Real-time PCR analysis
The proteome profiler results showed that there was different cytokine expression for each type of MSC including sICAM-1 (CD54), MCP-1 (CCL2), and SDF-1 (CXCL12). A real-time PCR analysis was used to quantify the sICAM-1 (CD54), MCP-1 (CCL2), and SDF-1 (CXCL12) transcripts. Their expression was normalized using the GAPDH housekeeping gene product as an endogenous reference. The primers and probes were designed for human sICAM-1, MCP-1, and SDF-1 using Primer Express 2.0 (Applied Biosystems, Foster City, CA, U.S.A.). sICAM-1 (CD54), MCP-1 (CCL2), and SDF-1 (CXCL12) mRNA levels were quantified using TaqMan Real-Time PCR with an ABI 7700 system (Applied Biosystems). Gene-specific probes and primer pairs for ICAM-1 (Assays-on-Demand, Hs00164932_m1; Applied Biosystems), MCP-1 (Assays-on-Demand, Hs00234140_m1; Applied Biosystems), and SDF-1 (Assays-on-Demand, Hs00171022_m1; Applied Biosystems) were used. For each probe/primer set, a standard curve was generated, which was confirmed to increase linearly with increasing amounts of cDNA. The amplification conditions were 2 min at 50℃, 10 min at 95℃, and a two-step cycle of 95℃ for 15 sec, and 60℃ for 60 sec for a total of 45 cycles.
Western blotting
Protein lysates were obtained with a buffer containing 50 mM HEPES (pH 7.5), 150 mM NaCl, 1.5 mM MgCl2, 1 mM EDTA, 10% glycerol, 1% Triton X-100, and a mixture of protease inhibitors (aprotinin, PMSF, and sodium orthovanadate). Equal amounts of total protein were resolved on a 12% SDS-polyacrylamide gel. The proteins were transferred to a nitrocellulose membrane. After blocking (TBS, 0.1% Tween-20) at 4℃ overnight, the membranes were incubated with primary antibodies of anti-mouse SDF-1, anti-mouse MCP-1, and anti-mouse sICAM-1 (R & D systems); all monoclonal antibodies were used at a dilution of 1:1,000 for 2 hr followed by incubation with secondary antibodies linked to HRP (Bio-Rad) and anti-rabbit GAPDH (dilution 1:2,000, Assay Designs, Inc., Ann Arbor Michigan, U.S.A.). Immunoreactive proteins were visualized by chemiluminescence using Super-SignalWest Dura Extended Duration Substrate (Pierce Chemical Co., Rockford, IL, U.S.A.). Fujifilm Luminescent Image Analyzer LAS-3000, with a charged-coupled device camera (Science Imaging Scandinavia AB), was used for imaging.
Statistics
Data are presented as the mean±SD. Data were analyzed with SPSS statistical software, version 12.0 (SPSS Inc, Chicago, IL, U.S.A.). The differences in cytokine concentrations among four groups of subjects were analyzed by the Kruskal-Wallis and Mann-Whitney tests. A probability of 0.05 was considered significant.
RESULTS
Isolation, culture, flow cytometry analysis and differentiation of placental MSCs
After enzyme digestion of the placental amnion and decidua, the cells were seeded into T-75 cell culture flasks at a confluence of 80%. The adherent cells with fibroblastic morphology were analyzed. The cells formed a monolayer of homogenous bipolar spindle-like cells with a whirlpool-like array in two to three days (Fig. 1), and these adherent cells could be readily expanded in vitro by successive cycles of trypsinization, seeding and culture every three days for three passages without visible morphologic change. The MSCs from term placental amnion and decidua were successfully isolated. We found no correlation between gestational age and the successful establishment of MSC cultures from the amnion and decidua.
We examined the surface marker profile of the amnion and decidua derived cell lines using fluorescence activated cell sorting (FACS). The phenotype of the MSCs derived from amnion was similar to that of MSCs derived from the decidua. These cells were positive for CD29, CD44, and CD73 but were negative for CD31, CD34, CD45, and HLA-DR (Fig. 2). CD 105 was positive in amnion MSC whereas CD 90 was positive in the decidua.
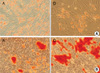
To estimate their potential to differentiate into several tissue lineages, the MSCs from the amnion and decidua were cultured in adipogenic, osteogenic, myogenic, and neurogenic medium. At the end of the induction period, the cells were differentiated into their respective tissues. The confirmation of differentiation was made by Oil Red O for adipogenic differentiation (Fig. 3A), Alizarin Red S staining for osteogenic differentiation (Fig. 3B), respectively. Culture expanded cells from the amnion and decidua were all able to differentiate into adipogenic and osteogenic lineages.
Cytokine array analysis
The conditioned medium, after 3-4 days of culture of the MSCs from CB, BM, and placenta (amnion, chorion), was assayed using the human cytokine array panel A (R & D systems) according to the manufacturer's instructions. We analyzed 36 cytokines at a time. The 36 cytokines included: C5a, CD40 Ligand, G-CSF, GM-CSF, GROα, I-309, sICAM-1, IFN-γ, IL-1α, IL-1β, IL-1ra, IL-2, IL-4, IL-5, IL-6, IL-8, IL-10, IL-12p70, IL-13, IL-16, IL-17, IL-17E, IL-23, IL-27, IL-32α, IP-10, I-TAC, MCP-1, MIF, MIP-1α, MIP-1β, Serpin E1, RANTES, SDF-1, TNFα, and sTREM-1.
CB MSCs secreted: MIF (GIF, DER6), IL-8 (CXCL8), Serpin E1 (PAI-1), GROα (CXCL1), and IL-6. BM MSCs secreted: MIF (GIF, DER6), IL-8, Serpin E1 (PAI-1), GROα (CXCL1), IL-6, MCP-1 (CCL2), and SDF-1 (CXCL12). Amnion MSCs secreted: GROα (CXCL1), sICAM-1 (CD54), IL-6, IL-8, MCP-1 (CCL2), MIF (GIF, DER6), and serpin E1. Decidua MSCs secreted: GROα (CXCL1), sICAM-1 (CD54), IL-6, IL-8, MIF (GIF, DER6), and Serpin E1.
Each of the MSCs expressed: MIF (GIF, DER6), IL-8 (CXCL8), Serpin E1 (PAI-1), GROα (CXCL1), and IL-6. However, MCP-1 (CCL2) was expressed only in the BM MSCs and amnion MSCs. sICAM-1 (CD54) was expressed in both the amnion and decidua MSCs. SDF-1 was expressed in only the BM MSCs (Fig. 4A). The relative expression level of the cytokines was calculated. The MCP-1 expression in the BM MSCs was higher than the expression in the amnion MSCs, whereas the expression of IL-6 in the CB MSCs was comparatively lower. GROα expression was higher in both the BM MSCs and amnion MSCs compared to the CB MSCs and the amnion MSCs (Fig. 4B).
The Detection of SDF-1 (CXCL12), MCP-1 (CCL2) and sICAM-1 (CD54) in each of the tissue-MSCs by RT-PCR
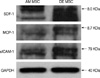
SDF-1 (CXCL12), MCP-1 (CCL2) and sICAM-1 (CD54) expression of human MSCs derived from three different origins were determined at the level of gene expression (n=8). Real time RT-PCR demonstrated that the human MSCs derived from the three different origins express cytokines differently. The level of mRNA was quantified and compared with a housekeeping gene (GAPDH). The higher expression of SDF-1 (CXCL 12) in BM MSCs was confirmed by RT-PCR (P=0.006). However, the RT-PCR did not confirm differences for the gene encoding MCP-1 (CCL2) among the four MSCs (P=0.252) derived from different tissues. The gene encoding sICAM-1 was found to be expressed at a higher level in the decidua MSCs (P=0.002) (Fig. 5). SDF-1, MCP-1, and sICAM-1 expression was confirmed by Western blot analysis in the amnion and decidua MSCs (n=2) (Fig. 6).
DISCUSSION
Mesenchymal stem cells are thought to have great therapeutic potential due to their capacity for self-renewal and multilineage differentiation (2, 13). They support hematopoiesis and enhance the engraftment of hematopoietic stem cells after co-transplantation (14, 15). Experimental and clinical data have demonstrated an immunoregulatory function of BM-derived MSCs (BM MSC) that may contribute to the reduction of graft-versus-host disease following hematopoietic stem cell transplantation (16, 17). Currently, the BM represents the major source of MSCs for cell therapy. However, aspiration of BM involves invasive procedures, and the frequency and differentiation potential of BM MSC decrease significantly with age (18). The search for alternative sources of MSCs, therefore, is of significant value. It has been reported that MSCs could be isolated from various tissues including periosteum, trabecular bone, adipose tissue, synovium, skeletal muscle, deciduous teeth, fetal pancreas, lung, liver, amniotic fluid, CB, and placental tissues (19, 20). Among these sources, CB and the placenta may be ideal sources due to their accessibility, painless donor procurement, promising sources for autologous cell therapy, and lower risk of viral contamination.
We isolated MSCs from the amnion and decidua. After isolation and culture, under specified conditions, typical MSC-like cells similar to the cells from the amnion and decidua were identified. These cells were characterized by the same shape and size, and the same monolayer appearance. In addition, the cells expressed CD29, CD31, CD34, CD44, CD45, HLA-DR, CD73, CD90, and CD105 by cytofluorimetry. Expression of the main marker genes of MSCs (CD29, CD44 and CD73) were positive and similar in all of the MSCs studied. (Data for BM MSCs and CB MSCs: not shown). Expression of CD31, CD34, CD45, and HLA-DR were negative. CD90 was positive only in the decidua-derived MSCs, whereas CD105 was positive only in the amnion-derived MSCs. The identity of MSCs from different sources has not been previously proven. The results of this study of placenta-derived MSCs may provide an attractive and rich source of MSCs.
Initially, cytokine analysis was performed with the proteome profiler™ to determine which cyotokines were expressed differently. Then, the cytokines were confirmed by PCR analysis. There was some discrepancy between the proteome profiler™ and the PCR analysis. For example, there was no significant difference in the expression of the gene encoding MCP-1 (CCL2) among the four differently derived MSCs by the PCR analysis. However, by the proteome profiler™ analysis, the amnion MSCs and BM MSCs were found to secrete MCP-1, whereas the CB MSCs and decidua MSCs did not secrete MCP-1. The gene encoding sICAM-1 was found to be expressed at a higher level in the decidua MSCs. The proteome profiler analysis, however, showed that the concentration of sICAM-1 was higher in the amnion MSCs compared to decidua MSCs. The manufacturing company recommended that the proteome profiler™ be used as a screening tool. Therefore, we depended on the results of the PCR analysis and Western blot to confirm the expression of SDF-1, MCP-1, and sICAM-1 in the MSCs from the amnion and the decidua.
Potian et al. (21) recently analyzed the cytokine profile of BM MSCs using the protein array technique. IL-6, IL-8, MCP-1, RANTES, GROα, INFγ, IL-1α, TGFβ, GM-CSF, angiogenin, and oncostatin M were constitutively expressed, and MIP-1α, IL-2, IL-4, IL-10, IL-12, and IL-13 were not expressed by the BM MSCs. The results of this study showed that the cytokine profile of the BM MSCs was very similar to that of the CB MSCs, with the exception that the CB MSCs expressed IL-12 but not G-CSF under serum-free conditions. Haynesworth et al. (22) reported that constitutively expressed cytokines in the growth phase include: IL-6, G-CSF, SCF, not detected in the growth medium of human BM derived MSCs. Both MSCs from the BM and CB abundantly produced IL-6, IL-8, and MCP-1.
The role that chemokines and their receptors play in the targeting of leukocytes to areas of inflammation, infection, or injury has been well characterized (23). As chemokine receptors are expressed on the cell surface of MSCs, and their stimulation has been shown to induce cell migration, it seems likely that they play a similar role in directing MSCs. MSCs have been shown to express a variety of chemokine receptors. The reported chemokine receptors of MSCs, however, have been inconsistent under similar isolation and culture conditions. This might be due to the heterogeneous nature of a typical MSC population that obscures the detection of a distinct receptor repertoire.
Our study findings showed that SDF-1 (CXCL12) was more highly expressed in the BM MSCs. SDF-1 is a small cytokine belonging to the chemokine family that is officially designated CXCL12. CXCL12 is strongly chemotactic for lymphocytes and is, therefore, an important hematopoietic growth factors for these cells (24-26). During embryogenesis it directs the migration of hematopoietic cells from the fetal liver to the BM. The receptor for this chemokine is CXCR4. This CXCL12-CXCR 4 interaction used to be considered exclusive, but recently it has been suggested that CXCL12 is also bound by the CXCR7 receptor (27, 28). SDF-1 could augment the mobilization, migration, recruitment, and entrapment of MSCs. SDF-1-CXCR4 interactions mediate the homing of MSCs. This suggests that SDF-1 plays an important role in the mobilization and homing of BM MSCs, although the signals required for this process have not been fully described.
The cytokine expression profile of placental MSCs remains poorly documented. We compared the cytokine expression of BM MSCs, CB MSCs, amnion MSCs, and decidua MSCs focusing on SDF-1 (CXCL12), MCP-1 (CCL2), and sICAM-1 based on the protein array assay. In this study, sICAM-1 was more highly expressed in decidua MSCs compared to the BM MSCs, CB MSCs and amnion MSCs. sICAM-1 represents a circulating form of ICAM-1 that is constitutively expressed or is inducible on the cell surface of different cell lines (29). sICAM-1 is a type of intercellular adhesion molecule continuously present in low concentrations in the membranes of leukocytes and endothelial cells. Upon cytokine stimulation, the concentrations of this molecule greatly increase. ICAM-1 can be induced by interleukin-1 (IL-1) and TNFα, and is expressed by the vascular endothelium, macrophages and lymphocytes (30). Although amnion MSCs and decidua MSCs are derived from the placenta, expression of sICAM-1 was found to be significantly lower in the amnion MSCs. Cytokine induction and signal transduction may be different in the amnion MSCs and the decidua MSCs. sICAM-1 is thought to play a more important role in the decidua MSCs than in the amnion MSCs with regard to cell recruitment. There was no significant difference in the expression of MCP-1 among the BM MSCs, CB MSCs, amnion MSCs, and the decidua MSCs.
In addition to their role in mediating cell migration, chemokines may also play important autocrine and paracrine roles. CXCL12 promotes the growth, survival, and development of MSCs (31). MSCs are known to be able to synthesize this chemokine, which may act in an autocrine manner via CXCR4. Chemokines are also recognized as primary inducers of integrin up-regulation following their interaction with their cell surface receptors and various downstream signaling events.
Integrins are known to mediate the firm adhesion of leukocytes to endothelial cells and play an important role in their transendothelial migration. It is likely that they play a similar role for the MSCs. MSCs are known to express various integrin molecules and their roles have begun to be elucidated. Pittenger et al. (2) reported the first study of integrin expression of MSCs and noted the presence of α1, α2, α3 aα, αν, β1, β3, and β4 along with the other adhesion molecules ICAM-1, ICAM-3, VCAM-1, ALCAM, and endoglin (CD105).
The results of the present study provide the characterization of cytokine expression of the MSCs from the BM, CB, amnion, and decidua. The cytokines play a role in migration of MSCs. The cytokine induction and signal transduction are important for migration of the MSCs. The characteristics of cytokine expression in MSCs derived from different tissues help with the understanding of the mechanisms of MSC migration. Further studies are required to better understand the interactions of these cytokines.



 XML Download
XML Download