

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The cornea is consisted of 3 different cell layers; epithelial cells, stromal fibroblasts (keratocytes), and endothelial cells. In the cornea, endothelial cells are the main target for the rejection in an allo-corneal transplantation, because fibroblasts are known to be less immunogenic than endothelial cells (1, 2).
In Asian countries, including Korea, cultural circumstances, including Confucianism, have caused a profound shortage of donor tissues for corneal transplantation. Therefore, the use of tissue from a xenogeneic source has been considered for a long-time by many Asian ophthalmologists. Recently, pig has been widely studied as a possible donor for xenotransplantation, because the pig's organ size as well as its anatomy and physiology make it an ideal substitute as a xenograft (3-5). The feasibility of porcine cornea as a xenograft has currently been evaluated (6). However, we observed that the xeno-corneal graft in a porcine to rat model showed different features from the murine or human allograft; the stromal fibroblasts, which are keratocytes, presented with more severe rejection than endothelial cells did (7). Hence, we wondered if the gene expression of porcine keratocytes would be different from that of corneal endothelial cells when it comes to the xeno-related rejection.
Since the first description of high-density DNA microarrays in 1995 (8), DNA microarrays have been widely used in genomics research. The long oligonucleotide microarray, which is composed of gene-specific oligonucleotides of 40-70 nt in length, spotted on glass slides, has become a powerful tool for globally detecting differential gene expression. Since the first-generation porcine oligonucleotide set, representing 13,297 cDNAs and expressed sequence tags (ESTs), has been designed by Qiagen-Operon for transcriptional profiling (9), we are now able to hybridize genes efficiently, using such porcine oligonucleotide set, to detect different expression levels between keratocytes and corneal endothelial cells.
Therefore, this study was undertaken to investigate the different gene expression of porcine keratocytes in xeno-related reaction in comparison to corneal endothelial cells, using cDNA microarray with porcine oligonucleotide set.
MATERIAL AND METHODS
The Ethnics Research Committee of the Seoul National University Hospital approved the protocol of the experiment.
Pig-to-rat orthotopic corneal transplantation
Twenty-one fresh porcine corneas, obtained within 6 hr of death, were used. To match the corneal thickness, the 500 µm anterior lamella was removed, using the Barron anterior chamber maintainer (Katena Products, Inc., Denville, NJ, U.S.A.) and crescent knife (Satin, Alcon surgical, Fort Worth, TX, U.S.A.). The remaining posterior lamella and endothelium was trephined from the inside with a 6 mm sized blade (Kai industries Co., ltd., Seki City, Japan), and the recipient rat cornea was also trephined carefully with a 4 mm sized blade (Kai industries Co., ltd), grasping the globe firmly with two fine tooth forceps until the immediate entrance into the anterior chamber. The graft was secured with 12 to 14 interrupted nylon sutures, and tarsorrhaphy was performed finally at the the lateral one-third of the lid margin. Antibiotics were instilled x3/day after surgery. No other medications were used. The rats were euthanized at 3, 7, 10, and 13 days after surgery, and the corneas were stained with hematoxylin-eosin staining.
Primary culture of porcine and human keratocytes
Ten porcine corneas were obtained from common adult pigs (approximately 80 kg in body weight). In accordance with the tenets of the Declaration of Helsinki and with properly informed consent, 10 human corneoscleral rims were obtained from the Northwest Lions Eye bank within 8 hr after the penetrating keratoplasty. The epithelial cells were removed, using a surgical blade, and the Descemet's membrane was mechanically peeled off. The remaining stroma was treated with 1.2 U/mL dipase I for 2 hr at 37℃, and 5 mL of type I collagenase was added and the tissues were shaken 3 times every 30 min. The harvested keratocytes were centrifuged at 1,200 rpm for 5 min, and the precipitate was inoculated into culture dish with DMEM:F12 (1:1) containing 10% FBS (fetal bovine serum; HyClone Laboratories, Utha, U.S.A.). The primary keratocytes were cultured at 37℃ in a carbon dioxide incubator for 1 to 2 weeks. The cells were passaged at 7-9 days before RNA isolation.
Primary culture of porcine endothelial cells
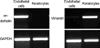
The Descemet's membranes were mechanically peeled off from the corneas of 50 common adult pigs. 0.05% trypsin and 0.01% EDTA were added to the tissues and the whole mixture was shaken 4 times every 20 min. The harvested endothelial cells were centrifuged at 1,200 rpm for 5 min and the precipitates were inoculated onto culture plate coated with collagen type IV. The resulting precipitate was submerged in M-199 medium (Cambrex, Charles City, IA, U.S.A.) containing 10% FBS. The primary endothelial cells were cultured at 37℃ in a carbon dioxide incubator for 1 to 2 weeks. The primary cells were passaged at 7-9 days before RNA isolation. Contamination was ruled out using flow cytometry after PE-rat anti- mouse CD14 (BD biosciences, Franklin Lakes, NJ, U.S.A.) and FITC-hamster anti-mouse CD3 (e-Bioscience, San diego, CA, U.S.A.) (1:100) staining for keratocytes and endothelial cells, respectively. We also confirmed that keratocytes and endothelial cells did not cross-contaminate with each other using the reverse transcriptase polymerase chain reaction (PCR) (RT-PCR) with the primer of vimentin (172 bp, 5'-AAGAGAACTTTGCCGTGGAA-3'; 3'-TCCAGCAGCTTCCTGTAGGT-5') and endothelin (329 bp, 5'-CCAAGGAGCTCCAGAAACAG-3': 3'-GGCAGAAACTCCAGCACTTC-5'
RNA isolation
Total RNA was extracted from the cells using Micro-to-Midi total RNA purification system (Life Technologies. Inc., Atlanta, GA, U.S.A.). Lysis buffer was added to the cells, and the cells were immediately homogenized using a Turrax homogenizer. The final product yielded 260/280 nm ratio of 1.8-2.0 and the purity was confirmed via gel electrophoresis. The concentration was determined based on 260 nm absorbance using a spectrophotometer.
Hybridization in microarray
A porcine platinum 13 K oligonucleotide microarray, which was developed in-house at GenoCheck Co., Ltd. (Ansan, Korea), was used for evaluation of the gene expression profiling. Control genes used in platinum 13 K oligonucleotide microarray chips were as follows: Arabidopsis thaliana photosystem I chlorophyll a/b-binding protein #X56062, RUB-ISCO activase #X14212, and ribulose-1,5-bisphophate carboxylase/oxygenase large subunit #U91966. Oligonucleotide microarray chips were prepared according to Zhao SH's updates of the genes (10). A total of 13,610 spots including control genes were spotted on platinum 13 K oligonucleotide microarray chips which were composed of 24 blocks with 568 genes each. We spotted and analyzed four different kinds of spike control samples on each block to confirm the quality of the chips and normalization of data. Oligonucleotide probes were resuspended in spotting buffer kit (GenoCheck Co., Ltd.) and spotted onto CMT-GAPS II slides (Corning Inc., Corning, NY, U.S.A.).
For microarray hybridization, total RNA of corneal endothelial cells was pooled and used for hybridization, and cDNAs were made from RNA sample via reverse transcription and labeled to produce the fluorescent-labeled cDNAs. Each total RNA sample (30 µg) was labeled with Cyanine3 (Cy3) or Cyanine5 (Cy5)-conjugated dCTP (NEN Life Science Product Inc. Boston, MA, U.S.A.) by a reverse transcription reaction using reverse transcriptase (Invitrogen, Carlsbad, CA, U.S.A.). The fluorescent-labeled cDNAs were mixed and simultaneously hybridized to the oligonucleotide microarray chip.
The oligonucleotide chips were scanned with an Axon 4000B Scanner (Axon Instruments, Foster City, CA, U.S.A.) using laser excitation of 2 fluorescence at 532 nm and 635 nm in wavelength for the Cy3 and Cy5 labels, respectively. The scanned images were analyzed with the software program GenePix Pro 5.1 (Axon Instruments, Inc., San Francisco, CA, U.S.A.) and GeneSpring GX 7.3.1 (Silicon Genetics, Santa Clara, CA, U.S.A.).
The raw intensity data were globally normalized by an intensity dependent on the LOESS (locally weighted regression scatter plot smoothing) method, and then was normalized by the with-print-tip group normalization method for each print-tip. The results were presented as M vs A scatter plots. Twenty four tips were used for making 13 K oligonucleotide microarray. Box plot normalization was performed to reduce variations of local background intensity caused by multiple pins (9). Over two-fold changed genes were selected for further analysis (M≥1 and M≥-1; M=log2 [red Cy5 intensity/green Cy3 intensity] ratio).
Real-time quantitative PCR
In order to validate the results from the microarray analysis, we selected four genes, which are C1R, CCL2, CXCL6 and HLA-A, and compared their gene expression in porcine keratocytes and corneal endothelial cells as measured by real-time PCR. Real-time quantitative PCR was performed in triplicate in 384-well plates. A 384-well high-throughput analysis was performed by using the ABI Prism 7900 Sequence Detection System (PE Applied Biosystems, Weiterstadt, Germany) and white colored 384-well plates (ABgene, Hamburg, Germany) for intensification of the fluorescent signals by a factor of three. The system operates using a thermal cycler and a laser that is directed via fiber optics to each of 384 sample wells. The fluorescence emission from each sample is collected by a charge-coupled device-camera and the quantitative data were analyzed using the Sequence Detection System software (SDS version 2.0, PE Applied Biosystems). Reaction mixtures contained 10 pM/µL of each primer and 2X SYBR Green PCR Master Mix (PE Applied Biosystems), which includes the HotStarTaqt DNA-Polymerase in an optimized buffer, the dNTP mix (with dUTP additive), the SYBRs Green I fluorescent dye, and ROX dye as a passive reference. Each of the 384-well real-time quantitative PCR plates included serial dilutions (1, 1/2 1/4, 1/8, and 1/16) of cDNA, which were used to generate relative standard curves for genes. All primers (Table 1) were amplified using the same conditions. Thermal cycling conditions 50℃ for 2 min and 95℃ for 10 min followed by 40 cycles of 95℃ for 30 sec and 60℃ for 30 sec, and 72℃ for 30 sec.
In order to exclude the presence of unspecific products, a melting curve analysis of products was performed routinely after finishing amplification by a high-resolution data collection during an incremental temperature increase from 60℃ to 95℃ with a ramp rate of 0.21℃/sec. We then converted real-time PCR cycle numbers to gene amounts (ng) on the basis of the equation. The real-time PCR analysis was performed on an Applied Biosystems Prism 7900 Sequence Detection System (PE Applied Biosystems).
RESULTS
Histology of rejected corneal xenografts in a pig-to-rat transplantation
The hematoxylin-eosin staining of cornea with time after porcine to rat corneal transplantation showed massive infiltration of neutrophils and monocytes into the stroma at the early stage (Fig. 1). Severe inflammation and rejection against stromal fibroblasts were observed.
Different gene expression in porcine keratocytes and corneal endothelial cells on microarray
After we confirmed that cultured keratocytes and endothelial cells did not cross-contaminate with each other using the reverse transcriptase PCR (RT-PCR) (Fig. 2), we used cDNA microarray to detect different expression levels between keratocytes and corneal endothelial cells. Hybridization image of the genes in keratocytes versus those in endothelial cells using a platinum pig 13 K biochip is presented in Fig. 3. Data intensity was normalized via M vs A scatter plot. Fig. 4 shows a log intensity signal ratio of the gene expression in porcine keratocytes versus that in porcine endothelial cells.
Up-regulated expression more than 2 folds was found in 1,162 genes of porcine keratocytes. Table 2 shows the genes involved in inflammation or immune reaction among the ones upregulated in porcine keratocytes than in porcine corneal endothelial cells. Of those immune-associated genes, whose signal intensity in porcine keratocytes was higher than that in porcine endothelial cells, the genes expressed more highly in porcine keratocytes than in human keratocytes were presented in Table 3; SEMA3C, CCL2, CXCL6, F3, HLA-A, CD97, IFI30, C1R, G1P3.
Validation of genes by real-time PCR
Real-time PCR showed that the expression of C1R, CCL2, and HLA-A was higher in porcine keratocytes, compared to that in porcine corneal endothelial cells (Fig. 5).
DISCUSSION
Allograft rejection in the corneal transplantation is mainly an endothelial rejection, and the stromal rejection against fibroblasts is very rare in human or murine allograft (2). Surprisingly, however, we observed severe inflammation and rejection against stromal fibroblasts in an orthotopic porcine to rat corneal transplantation (Fig. 5). In addition, in a mixed reaction with human sera or human PBMC (7), we found a higher susceptibility of porcine keratocytes to an immune-mediated damage than corneal endothelial cells. This is the reason why we analyzed and compared the gene expression of porcine keratocytes and porcine endothelial cells in this study. We tried to elucidate major factors involved in the xeno-related stromal rejection.
First, we found 20 genes having high signal intensity ratios of porcine keratocytes/endothelial cells among the immune-related genes. Next, we compared the expression of those candidate genes with their expression in human keratocytes and sorted out highly expressed genes, because they might be involved in the development of porcine stromal inflammation or rejection, which was not observed in stromal keratocytes of human allograft. As a result, target genes turned out to be SEMA3C, CCL2, CXCL6, F3, HLA-A, CD97, IFI30, C1R, and G1P3. When we compared the gene expression of C1R, CCL2, CXCL6, and HLA-A between porcine keratocytes and porcine endothelial cells using the realtime PCR, C1R, CCL2, and HLA-A were highly expressed in porcine keratocytes. Notably, the level of C1R gene in porcine keratocytes was as almost 16 times as that in porcine endothelial cells.
C1R is a highly specific serine protease that initiates the classical pathway of complement activation. A report presents elevated levels of C1rs-C1inh complex in tears after the corneal transplantation, suggesting that the classical pathway of the complements may be activated in the early postoperative period of the corneal transplantation (11). Especially, complement-associated hyperacute- or acute- rejection is regarded as a very important rejection mechanism in xenograft of vascular organs such as kidney or heart. However, the effect of complements on rejection has been overlooked in corneal allograft because the concentration of complements in aqueous humor is lower than that in serum. Moreover, complements cannot reach the corneal stroma due to avascularity of corneal stroma especially in normal quiet cornea. Usually, complement proteins are mainly synthesized by hepatocytes but are also secreted by tissue macrophages and blood monocytes (12). The complements are also known to be secreted by keratocytes (corneal fibroblasts) (13). High secretion of complements by keratocytes is likely to be crucial in corneal xenograft, as well as the rapid infiltration of innate immune cells, that synthesize complements, may also contribute to increase the concentration of complements in the stroma. There is a report showing keratocyte apoptosis in an orthotopic human to cat corneal xeno-transplantation (14). The percentage of TUNEL-positive cells was higher in the stromal keratocytes than in the endothelial cells, which is consistent with our results. Taken all together, increased expression of C1R gene in porcine keratocytes provides a plausible evidence on the possible complement-mediated stromal rejection of the porcine corneal xenograft.
CCL2 (MCP-1) displays a chemotactic activity for monocytes and basophils, but not for neutrophils or eosinophils. Many studies demonstrate the increase in MCP-1 and subsequent macrophage infiltration to the pig islet xenograft (15-17) and pig heart xenograft (18). Likewise, our in-vivo porcine to rat corneal transplantation showed early neutrophil migration and subsequent monocytes/macrophages infiltration (Fig. 5).
HLA-A or MHC Class I molecule is a major antigen to the immune system, and is easily recognized by CD8+ T-lymphocytes. Our porcine xenograft demonstrated high CD8+ infiltration as early as 7 days after transplantation (unpublished data), and this finding seems to be associated with the increase of HLA-A gene expression in this study.
Recruitment of neutrophil to inflammatory sites is mediated by two related receptors: CXCR1 and CXCR2. Both receptors share two ligands, interleukin-8 (CXCL8) and GCP-2 (CXCL6). The role of CXCL6 in inflammation to facilitate neutrophil infiltration is well established (19), while its involvement in xeno-related rejection is uncertain. The porcine islet graft presents mostly monocytes/macrophages and T cells migration, but not neutrophils (15, 16). We found relative increasing expression of CXCL6 gene in porcine keratocytes, which is 16 and 8 folds higher than in human keratocytes and porcine endothelial cells, respectively, whereas real time PCR revealed expression of RNA did not. We do not know exact roles of CXCL6 right now, because we could not perform the real time PCR between porcine keratocytes and human keratocytes, due to technical difficulties that the porcine primers are different from human's. If the expression would be higher in porcine keratocytes than in humans, there may be a still possibility of CXCL6 contribution to xeno-associated stromal rejection on the contrary in human.
Even if we did not perform the real-time PCR analysis for SEMA3C, F3, CD97, IFI30, C1R, and G1P3 genes, it is possible that high expression of those genes on the microarray analysis might be related to the stromal rejection observed in a porcine to rat corneal xenograft.
SEMA3C is known to promote glomerular endothelial cell proliferation, adhesion, directional migration, and tube formation in vitro by stimulating integrin phosphorylation and VEGF120 secretion. The functional role of SEMA 3C in rejection has not been reported yet.
Tissue factor (F3, thromboplastin) is the cell surface receptor for the serine protease factor VIIa. The best known function of F3 is its role in blood coagulation. The complex of F3 with factor VIIa catalyzes the inactive protease factor X into the active protease factor Xa. Together with factor VII, tissue factor forms an extrinsic pathway of coagulation. Their role in the humoral rejection of a xenograft is well-established (20, 21). However, the involvement of F3 in the corneal rejection is not clear, because there are no vessels in the stroma of the cornea.
CD97 has an extended extracellular region with several N-terminal epidermal growth factor (EGF)-like domains, which mediate binding to its cellular ligand and decay accelerating factor (DAF, CD55). The presence of structural features characteristic of extracellular matrix proteins and transmembrane proteins suggests that this protein is a receptor involved in both cell adhesion and signaling processes early after leukocyte activation. However, little is known about the physiological function of CD97. In fact, a report suggests that absence of CD97 does not impede migration of granulocytes to the sites of local tissue (22).
The IFI30 protein is a lysosomal thiol reductase that can reduce protein disulfide bonds at low pH. The enzyme is expressed constitutively in the antigen-presenting cells and can be induced by gamma-interferon in other cell types. This enzyme has an important role in MHC class II-restricted antigen processing by unfolding the antigenic proteins in preparation for their proteolytic cleavage and presentation of resulting peptides by MHC class II. It has also been shown to suppress T cell activation (23). Their critical function in immune regulation remains still controversial, and the role as an enhancer of granulocyte infiltration is not known, either.
GIP3 protein interactions possibly play a pivotal role in the regulation of apoptosis (24). However, the detailed function of GIP3 protein as well as an immune modulation, still needs to be analyzed.
Our study has major limitations. First, we only evaluated the gene expression without protein data, because antibodies for the porcine functional proteins are not available. Moreover, we mainly analyzed the genes that were associated with neutrophil accumulation, because we observed massive neutrophil infiltration in the early stage after pig-to-rat corneal xenotransplantation. Other genes than those analyzed in this study might be involved in the xenorejection process observed in our model. Second, we used cultivated primary cells to detect different levels of gene expression. It would be more valuable if we use cells collected from rejected corneal xenografts rather than cells cultivated and collected from fresh cornea, because cells can be transformed during cultivation procedure, and it might not precisely reflect the pattern of gene expression during the early rejection period. Third, we analyzed different gene expression between human and porcine corneal cells based on the observation in a pig-to-rat model. Nevertheless, we still believe that the data provided in this study are worthy of understanding the rejection mechanism in porcine to rat xenotransplantation and further predicting the immune reaction in porcine to human xenotransplantation.
In conclusion, based on the known evidence-based function, the increased expression of C1R, CCL2, and HLA-A appears to be responsible for the stromal rejection of keratocytes in xeno-related corneal rejection. It can be mediated by the recruitment of acute inflammatory cells or possibly due to the complement-associated killing. The roles of other genes, which showed increased ratio, in the stromal rejection remain to be further investigated.







 XML Download
XML Download