

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Partial trisomy 1q syndrome is a rare chromosomal abnormality, arising in most cases from a parental balanced translocation with partial monosomy of other autosomes. De novo translocation, duplication or insertion has also been reported (1, 2). To date, two major partial trisomy 1q syndromes with regard to the breakpoint localization have been described, 1q32-qter and 1q42-qter (3), with the former described as "proximal partial trisomy" and the latter as "distal partial trisomy 1q" (2). Although several cases of pure proximal partial trisomy 1q have been reported (4, 5), distal partial trisomy 1q syndromes are frequently accompanied by other chromosome aberrations, making the definition of a phenotype difficult. However, individuals with partial trisomy 1q42-qter, combined with small deletions of telomeric segments of various autosomes, showed a milder phenotype than those with partial trisomy 1q32-qter (2, 6-8).
We have utilized array comparative genome hybridization (CGH) to delineate a nearly pure trisomy 1q41-qter that showed a trisomy 1q42-like phenotype, and we characterized the breakpoints in detail.
CASE REPORT
The proband was a 6-month-old male child born to a non-consanguineous, phenotypically normal couple. He was born at 39 weeks of gestation by cesarian section with a birth weight of 2,820 grams. At the age of 6 months, his weight was 5.6 kg (<3rd percentile), his height was 63 cm (<3rd percentile), and his head circumference was 46 cm (90th percentile). He showed poor growth and development, including absence of social smiling and maternal recognition at the age of 12 months. Mild dysmorphic features were present in Table 1. Echocardiography, electromyography, hematological and biochemical tests for renal and liver function showed no abnormalities, and screening for inborn errors of metabolism was normal.
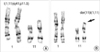
Conventional cytogenetic analysis of the proband revealed an abnormal male karyotype, 46,XY,der(11)t(1;11)(q41;p15.5) (Fig. 1). The proband's mother and sister had the same balanced karyotype, 46,XX,t(1;11)(q41;p15.5) with a normal phenotype.
Further characterization of the size and boundaries of the cytogenetic abnormalities of the proband was carried out by array CGH. Genomic DNA was extracted from peripheral blood using QIAamp DNA Micro kit (Qiagen, Hilden, Germany). The arrays (MacArray Karyo4000 from Macrogen, Seoul, Korea) were consisted of 4,000 human bacterial artificial chromosomes (BACs), spaced approximately 1 Mb across the whole genome, with each BAC clone spotted in duplicate. Labeling and hybridization were performed as described (9). Arrays were scanned on a GENEPIX4200A two-color fluorescent scanner (Axon Instruments, Union City, CA, U.S.A.). The log2-transformed fluorescence ratios were calculated from median intensity values, after subtracting for background, and normalized according to the intensity normalization method. Chromosomal aberrations were categorized as a gain or loss when the normalized log2 transformed fluorescence ratio was outside the range 0.25 to -0.25. These two threshold values were chosen by selecting a 3SD (standard deviation) value calculated from 30 normal male to normal female hybridization experiments.
The array CGH for the proband revealed that the breakpoint of a contiguous duplication on the long arm of chromosome 1 was located in 1q41 band (Fig. 2). We also observed two BAC clone deletions on the short arm (11p15.5) of chromosome 11 (Fig. 2).
To confirm the gain and loss predicted by the array CGH, fluorescence in situ hybridization (FISH) analysis was performed and the array CGH analysis of proband's mother was also done in order to exclude the copy- number variations showed by the array CGH results of the proband. BAC clones (Macrogen, Seoul, Korea) for FISH analysis included BAC_4527 (position start and end number; 219935200-220017617) proximal to, and BAC_7952 (220560043-220660043) distal to the break region of 1q41 predicted by the array CGH. BAC_1341 (202674-299580) and BAC_5807 (257732-338908) were used for the deletion of 11p15.5 predicted by the array CGH. Additionally, FISH with BAC_4069 (238256420-238340569) was carried out to confirm whether it is a continuous duplication or two separated duplications around 1q43 that showed no duplication pattern by the array CGH. At least twenty metaphase and 200 interphase preparations were scored for each hybridization.
FISH analysis confirmed the breakpoint of a contiguous duplication on the long arm of chromosome 1 was located between BAC_4527 and BAC_7952 (more proximal to the BAC_4555) (Fig. 3A). Additionally, the skipped duplication pattern by the array CGH around at 1q43 region was proved to be false by FISH with 1q43 probe (Fig. 3B). The deletion of 11p15.5 predicted by the array CGH was also confirmed by FISH (Fig. 3C).
DISCUSSION
The clinical phenotypes of partial trisomy 1q syndrome vary widely, due to the different breakpoints on chromosome 1 and the extent of the monosomic segment of the involved autosomes. Partial trisomy 1q syndrome can be classified according to breakpoint position as 1q32-qter or 1q42-qter (2, 3). Duplication of 1q42-qter with no other involved chromosome usually presents as a mild phenotype, which may include macrocephaly with wide fontanelles, flat nasal bridge, low-set ears, facial capillary nevi, growth retardation, and developmental delay (2, 8). Partial trisomy 1q32 syndrome, however, is usually characterized by more severe phenotypes, including urogenital anomalies and severe cardiac defects (3).
The patient described here, with partial trisomy 1q syndrome and a mild phenotype, showed phenotypic similarities to pure partial trisomy 1q42 syndrome, due to the minimal involvement of chromosome 11, in only the distal telemetric region (Table 1). The deleted region of 11p15.5 in this case showing two-clone deletions of BAC_1341 and BAC_5807 by the array CGH included following genes; RIC8A, SIRT3, PSMD13, COX8B, NALP6, ATHL1, MRPS24P1, NALP6, ATHL1, MRPS24P1, IFITM1, IFITM2, and IFITM3. None of these genes are known to be related to the clinical abnormal phenotypes. The critical genes for well-known genetic diseases such as hyperproinsulinemia (OMIM+176730), maturity-onset diabetes of the young, type I (OMIM #125850), Beckwith-Wiedemann syndrome (OMIM #130650) and hereditary adenocortical carcinoma (OMIM #202300) were located in much more proximal from the deleted region of this case and these genes were not involved.
The array CGH showed that the partial trisomy 1q syndrome in our patient had nearly the same breakpoint as the first partial trisomy 1q41-qter case reported. The marker D1S2439 in sub-band 1q41 was reported to be critical for the presence of urogenital abnormalities (2). This D1S2439 is located between BAC_4527 (219935200-220017617, the most distal marker; MFD255) and BAC_4555 (221038499-221109556, the most proximal marker SHGC-78488) of 1q41 of our case. The FISH probes made of BAC_4527 and BAC_7952 which included D1S2439 and BAC_4555 confirmed the breakpoint predicted by the array CGH and the urogenital abnormalities were not found in our case. Therefore, the breakpoint in a severe form of partial trisomy 1q syndrome (so called, proximal partial trisomy 1q syndrome) is expected to be located more proximal to D1S2439. However, with the exception of our case and that of Emberger et al., we could hardly find studies that had utilized molecular techniques such as FISH or array CGH to identify the exact breakpoint of distal trisomy 1q syndrome (6, 8).
To our knowledge, this is the first case of partial trisomy 1q41 syndrome in which whole genome array CGH was used to identify other submicroscopic deletions or duplications over the entire genome. Use of array CGH to define the breakpoint may be time-saving and cost-effective compared with using multiple FISH probes. The reproducibility and the accuracy of array CGH relative to FISH and real-time PCR methods have been validated (10, 11). Array-based CGH has become a powerful method to detect and analyze genomic imbalances that are not detected at the level of high resolution banded karyotype analysis.



 XML Download
XML Download