

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Chromosomal aberrations can be a hallmark of cancer, and molecular cytogenetic analyses have a considerable potential as diagnostic adjuncts. Several chromosomal and molecular abnormalities have been identified in non-small cell lung carcinomas (NSCLC) (1, 2). Although early-stage carcinomas have been found to have fewer molecular alterations than advanced-stage carcinomas, such early molecular changes might be used as molecular screening tools for the early detection of NSCLC in high-risk patient populations.
Fluorescence in situ hybridization (FISH) has been used successfully for targeted analysis of numerous chromosomal abnormalities in interphase nuclei of various types of solid tumors (3). Despite the high frequency of aneusomy in lung cancer, FISH has not yet been fully exploited for early detection and monitoring of this tumor type, in part because of the unavailability of validated probes specifically for lung carcinoma. We used the recently developed multi-target FISH probes, LaVysion (Vysis, Downers Grove, IL, U.S.A.), for simultaneous analysis of chromosome 6 and the 5p15.2, 7p12 (EGFR gene), and 8q24.12-q24.13 (MYC gene) loci. This made it possible to simultaneously screen copy number changes for four distinct DNA sequences.
Genomic alterations of regions on chromosome arm 5p have been observed frequently in lung cancers (4); however, copy number alteration has rarely been described in early-stage lung cancer. Understanding chromosomal alterations in early-stage lung cancer led to the discovery of candidate genes and will also offer an insight into its pathogenesis. Moreover, classical cytogenetics (5), interphase FISH (6-8), and comparative genomic hybridization (9, 10), have shown gains in chromosomes 6, 7, and 8 in approximately 50% of the NSCLC, which suggests that chromosomes 5, 6, 7, and 8 might be suitable targets for diagnostic FISH probes.
In this study, a multi-target FISH assay was performed to investigate the correlation between chromosomal alterations in each probe and clinicopathological variables, and to clarify the potential of the multi-target FISH assay in patients with stage Ia-IIIa NSCLC.
MATERIALS AND METHODS
Patients and sample preparation
Touch imprints were prepared from 37 NSCLC at the time of surgical resection. At the same time, fifteen imprints of normal areas were obtained from the lobected tissue of the controls under the guidance of a pathologist. All subjects had a lung mass that was operable. In addition, none of them had undergone chemotherapy or irradiation. Imprints were obtained according to a slightly modified version of the protocol described by Varella-Garcia et al. (11). The freshly resected tumor was imprinted on silanized slides. After washing, the specimens were fixed in a 3:1 solution of methanol:glacial acetic acid and were stored at -20℃ until analysis. The charts of all patients were reviewed for any significant medical history. Permission to perform the study was obtained by the institutional review board (IRB) at Chungnam University Hospital, Daejeon, Korea.
Fluorescence in situ hybridization
A LaVysion kit (Vysis, Downers Grove) was used for FISH. The probe set included one centromeric sequence (6p11.1-q11) labeled with SpectrumAqua and three single copy DNA sequences. These sequences recognize 450-kb sequences mapped at 5p15.2 (SpectrumGreen), 7p12 (including the EGFR gene, SpectrumRed), and 8q24.12-q24.13 (including the MYC gene, SpectrumGold).
The laboratory process was performed according to the recommendations of the manufacturer with minor modifications. In brief, the slides were denatured by incubation with formamide (70% in 2×SSC) at 73℃ for 5 min in a water bath. The slides were then dehydrated through a graded ethanol system (70% for 1 min, 85% for 1 min, and 100% for 1 min). A hybridization solution (10 µL) was applied to each slide, which was then cover-slipped and sealed with rubber cement. After incubation for over 16 hrs at 37℃ in a humidified chamber, the slides were washed with 0.4×SSC/0.3% NP-40 for 2 min at 73℃. The slides were placed in 2×SSC/0.1% NP-40 for 1 min at room temperature. A 4, 6-diamidino-2-phenylindole-antifade solution (10 µL) was then applied to each spot, which was subsequently cover-slipped.
The slides were observed under a fluorescence microscope connected to a cooled, charge-coupled device camera and an image analyzer system, CytoVision (Applied Imaging, Ltd., Newcastle, United Kingdom). Two observers were independently involved in the FISH analysis, and investigators responsible for the FISH analysis were blinded to the status of specimens, regarding all variables, except for the date of specimen collection. The score was calculated as an average between two observers' scores.
The signals were scored on a cell-by-cell basis after hybridization and washing. The slides were assessed by scanning for morphologically abnormal cells with nuclear enlargement, irregular nuclear contour, patchy or lighter nuclear DAPI staining, and cell clusters, all of which were indicative of malignancy (12, 13). A normal cell contains two red, two gold, two green, and two aqua signals. A minimum of 25 morphologically abnormal cells were assessed, or the entire sample was screened. Slides that contained fewer than six cells were considered to be inadequate for evaluation. Overlapping cells or cells with blurred signals were not assessed. Signals that were located very close to each other were interpreted as split signals and were counted as one signal. Slides showing suspicious signals were re-examined by another technician to verify the results. The test results were defined based on previous studies (7, 14, 15). The identification of 6 or more cells with polysomy, defined as a gain of 2 or more chromosomes in a cell, defines cancer positivity in this study.
Statistical analysis
We utilized several techniques for statistical analysis to verify our results. Possible correlations between clinicopathological variables (cancer type and lymph node metastasis) and the mean chromosome copy number were evaluated via independent t-tests. We employed one-way ANOVA (analysis of variance) in our comparisons of the differences in the mean chromosome copy number between TNM (tumor-node-metastasis) tumor stages. p values <0.05 were considered significant.
RESULTS
The overall sensitivity of FISH in lung cancer
FISH was performed to detect genetic alterations of chromosomes 5p15.2, 6p11.1-q11, 7p12, and 8q24.12-q24.13 (LaVysion Test), which showed very high sensitivity in detecting stage Ia to IIIa non-small cell lung cancers. A summary of clinicopathological data of 37 non-small cell lung cancer patients are shown in Table 1. When the aneusomy rate was defined as the percentage of carcinomas with aneusomy for at least two DNA targets, the aneusomy rate with the LaVysion probe was 92.6% (35/37). Abnormalities of at least one of the chromosomes were observed in all cases (37/37), and 12 tumors (32.4%) showed polysomy of all four chromosomes. A gain in 5p15.2 (27/37=73.0%) was the most recurrent finding, followed by gains in 7p12 (24/37=64.9%), 8q24.12-q24.13 (21/37=56.8%), and 6p11.1-q11 (15/37=40.5%). All 37 tumor imprints showed good morphology. We found no false-positive cases among the 15 normal control specimens (data not shown). Fig. 1 shows representative examples of the positive FISH results from patients with lung cancer.
The relationship between each probe and the clinicopathological variables
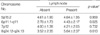
In order to further clarify the chromosomal alterations in lung cancer, we explored the statistical relationship between the mean chromosome copy number and clinicopathological variables in each probe. The mean chromosome copy numbers, in descending order, were 5p15.2 (4.62), 8q24.12-q24.13 (4.32), 7p12 (4.08), and 6p11.1-q11 (3.41) (Table 2). The 5p15.2 region was the most frequently involved, and was observed in 22 early-stage (I+IIa) lung cancers. The mean chromosome copy number (4.63) in 16 stage I cancers was considerably higher than in other regions, followed by 7p12 (3.94), 8q24.12-q24.13 (3.13), and 6p11.1-q11 (2.50). This region was detected in 81.3% of 16 stage I cancers. Gains of 6p11.1-q11 and 8q24.12-q24.13 were observed in 61.5% (8 of 13) and 84.6% (11 of 13) of stage IIIa cancers, respectively, while lower frequencies were seen in 25.0% (4 of 16) and 31.3% (5 of 16) of stage I cancers, respectively (Table 2). Furthermore, the mean chromosome copy number exhibited a tendency to increase directly with TNM stage, and a significant difference was shown (p=0.024 and p=0.017, respectively) (Table 2).
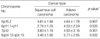
The frequencies of gene amplification and/or hyperaneusomy at 6p11.1-q11 and 8q24.12-q24.13 were statistically significantly correlated with specific clinicopathological parameters (p<0.05); TNM tumor stage (I [a+b], II [a+b], IIIa), lymph node status (absent or present), and histological type (adenocarcinoma or squamous cell carcinoma (Table 2-4). Tumors with lymph node metastasis were found to show higher frequencies than tumors without lymph node metastasis (p=0.025 and p=0.013, respectively) (Table 3). The correlation was significantly higher in adenocarcinoma compared to squamous cell carcinoma (p=0.030 and p=0.020, respectively), whereas the number of imbalances of 5p15.2 was similar in squamous cell carcinoma and adenocarcinoma (Table 4). Gains of 8q24.12-q24.13 were seen in 71.4% (10/14) of adenocarcinoma compared to 47.8% (11/23) of squamous cell carcinoma, while gains of 6p11.1-q11were detected in 64.3% (9/14) of adenocarcinoma but in only 26.1% (6/23) of squamous cell carcinoma.
DISCUSSION
Earlier identification of cancer cells will lead to a greater understanding of the chromosomal alterations that occur, and will allow for a more complete insight into the pathogenesis and progression of lung cancer. However, no efficient or accurate screening methods have yet been discovered for early detection of lung carcinoma.
FISH can easily detect the number of specific chromosomes or certain gene loci in histologic or cytologic tumor specimens, and may be conducted at a very early stage or even at the precancerous stage. The recent development of multi-target LaVysion FISH probes made it possible to simultaneously screen copy number changes for four distinct DNA sequences; in each of our samples, an abnormality of at least one of the chromosomes was observed and 12 tumors (32.4%) showed polysomy of all four chromosomes.
Classical cytogenetics (5), interphase FISH (6-8) and comparative genomic hybridization (9, 10) show gains of chromosomes 6, 7, and 8 in approximately 50% of NSCLC. A gain of 5p is also a very common recurrent abnormality in SCLC (9, 16), which suggests that chromosomes 5, 6, 7, and 8 might be suitable targets for diagnostic FISH probes. This suggestion was confirmed by the present study, which indicated a high level of sensitivity (35/37=92.6%) of the four-probe LaVysion assay in lung cancer, and specificity was 100%.
In the present study, a gain of 5p15.2 was the most frequent alteration (27/37=73.0%), and the mean chromosome copy number in stage I was considerably higher than in other regions. Garnis et al. (4) suggested that gains on 5p indicate early events in lung cancer, and that the amplification of TRIO and GDNF genes at 5p were some of the most dominant phenomena in NSCLC. Several molecular cytogenetic studies have shown that chromosomal aberrations occur on the short arm of chromosome 5 in all major lung tumor types, and other studies have reported gains on 5p to be among the most frequent alterations in SCLC (7, 16). These findings suggest that gene amplification in the 5p region is one of the most consistent alterations in early-stage lung cancer and the amplification of this region is a major susceptibility locus in lung cancer.
Chromosome 7 was shown to be an early marker of tumorigenesis in several types of cancers; polysomies of this chromosome have been demonstrated in hyperplastic and dysplastic epithelium by FISH (17). We detected 64.9% polysomy at 7p12, including the epidermal growth factor receptor, EGFR, and IL-6 genes, which are located on chromosome 7p and are overexpressed in NSCLS (18). Zojer N et al. (6), demonstrated that aberrations of chromosome 7 occur early in the development of lung cancer and that the aneuploidy rate of chromosome 7 was 100% in primary lung cancer. EGFR overexpression has also been demonstrated in premalignant bronchial epithelium, which suggests that EGFR plays an important role in lung carcinogenesis (18)
A gain at 8q was the most frequently encountered aberration in 13 stage IIIa cancers (84.6%), with a minimal region occurring at 8q24.12-q24.13, the region in which the c-MYC oncogene is located. Several studies suggest that c-MYC amplification or overexpression is associated with a worse prognosis (19). Pei et al. (20) suggested that a gain of 8q was associated with later stage, higher grade tumors. Kubokura et al. (19) also suggested that the rate of chromosome 8 aberration is correlated with longevity of survival rate. Amplification of the MYC family of proto-oncogenes (MYCC, MYCN, and MYCL1) in human lung cancers has been well characterized (21). MYCC is amplified in both SCLC and NSCLC, while MYCN and MYCL1 amplifications are usually limited to SCLC (21). In this study, the mean chromosome copy number at 8q24.12-q24.13 was significantly correlated with the stage of TNM tumor (p=0.017). Tumors with lymph node metastasis were found to occur at a higher frequency than tumors without lymph node metastasis (p=0.013). We considered the rate of chromosome 8 aberration to be an additional prognostic factor of patients with NSCLC.
In this study, we detected 40.5% polysomy on chromosomes 6p11.1-q11. Juan et al. (22) suggested that amplification in 6p is the most prominent finding in NSCLC. In the present study, amplification of chromosome 6 was significantly correlated with advanced clinical stage and lymph node metastasis. These findings suggest that gene amplification of chromosome 6 is strongly associated with the development, progression, and metastasis of tumors.
Squamous cell carcinoma (SqC) and adenocarcinoma (AdC) are the two most common subtypes of NSCLC. A growing line of evidence suggests that the SqC and AdC subtypes progress through different carcinogenic pathways, but the genetic aberrations promoting such differences remain unclear. In the present study, the frequencies of gene amplification and/or hyperaneusomy at 6p11.1-q11 and 8q24.12-q24.13 were significantly higher in AdC. These data suggest that genes located in these chromosomal regions may be associated with phenotypic properties that differentiate SqCs from AdCs.
In conclusion, our findings suggest that gains at 5p15.2 might be early events of NSCLC, and gene amplification in this region might be strongly associated with the development. Additionally, genes located at 6p11.1-q11 and at 8q24.12-q24.13 may be associated with tumor aggressiveness and could represent markers of lung cancer progression.



 XML Download
XML Download