

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Lung cancer is the most prevalent and lethal cancer worldwide, and has been reported in many nations, including the Republic of Korea and many Western countries (1). The high mortality rate attributed to lung cancer may be the result of induced multi-drug resistance and a high frequency of metastatic occurrence. Despite aggressive treatment with surgery, radiation, and chemotherapy, survival rates for lung cancer patients remain low (2). New approaches to the treatment of lung cancer are urgently needed. Immunotherapy may represent one such approach, and several different strategies for lung cancer immunotherapy have recently been developed (3).
The goal of cancer immunotherapy is to augment the weakened host immune response against tumors using specific and/or nonspecific immune stimulants. Treatment may include the administration of cytokines, tumor-specific antibodies, and T-cells (3-5). Toxoplasma gondii is an obligate intracellular parasite; latent infections with T. gondii are common in human populations throughout the world (6). It has been reported that the development of spontaneous mammary tumors, leukemia, and 20-methylcholanthrene-autoinduced tumors is suppressed in animal models following injection of Toxoplasma antigen or viable parasites (7, 8). Both Toxoplasma infection and the cell-free parasite extract are able to reverse the multi-drug resistance of mouse lymphoma and human gastric cancers in vitro (9). Moreover, potent antitumor effects are induced by intralesional injection with formalin-fixed T. gondii organisms in Lewis lung carcinoma (LLC) in Toxoplasma-infected mice (10, 11).
These reports suggest that T. gondii is a powerful agent for cancer immunotherapy, and is useful as a stimulant of the cellular immune responses. Recently, there were many reports that antitumor and antimetastatic actions are associated with inhibition of tumor-induced neovascularization (12-14). However, the immunologic mechanisms of the antitumor activity elicited by T. gondii in lung cancer, as well as the immunological characteristics of LLC, are not well documented. Also, the anti-angiogenic effects of T. gondii have not been determined in the LLC mouse model. In order to determine the antitumor and anti-angiogenic activities of T. gondii infection in LLC-bearing mice, C57BL/6 mice were injected with LLC cells alone or in combination with T. gondii. And then, we evaluated the survival rates, tumor size, histopathology, cell-mediated immune responses, and angiogenesis.
MATERIALS AND METHODS
Mice and parasite strains
Female inbred C57BL/6 mice were obtained from the Korea Research Institute of Bioscience and Biotechnology, Daejeon, Korea. All mice used were 6-8 weeks old and documented as specific pathogen-free animals. All animal studies were carried out in a pathogen-free barrier zone at the College of Medicine, Chungnam National University, in accordance with the procedures outlined in the Guide for the Care and Use of Laboratory Animals. Two strains of T. gondii were used; the RH strain was used to prepare Toxoplasma lysate antigen (TLA) and mice were orally infected with the Me49 strain to evaluate antitumor activity.
Lewis lung carcinoma (LLC) cell cultures and experimental groups
The LLC cell line was obtained from the American Type Culture Collection (ATCC, Rockville, MD, U.S.A.) and was cultured in a humidified atmosphere of 5% CO2 at 37℃ using Dulbecco's modified Eagle's medium (DMEM) (GibcoBRL Co., Grand Island, NY, U.S.A.) that contained 10% heat-inactivated fetal bovine serum (FBS; GibcoBRL), 2 mM glutamine, 100 U/mL penicillin, and 100 µg/mL streptomycin.
Mice were divided into seven experimental groups. Each group was composed of 50 mice; 10 mice to evaluate survival rates, 35 mice to evaluate immunologic characteristics, and 5 mice to check angiogenesis. The seven experimental groups included mice that were administered with: LLC cells (LLC-injected group); T. gondii (TG-injected group); both Toxoplasma parasites and LLC cells (TG/LLC-injected group); Quil-A only (QA-injected group) (Accurate Chemical and Scientific Co., Westbury, NY, U.S.A.); both LLC cells and Quil-A (LLC/QA-injected group); both T. gondii and Quil-A (TG/QA-injected group); and T. gondii, LLC cells and Quil-A (TG/LLC/QA-injected group). To calculate the survival rate for each group, the mice were examined each day for eight weeks following LLC cell injection or T. gondii infection. To evaluate the immunological mechanisms of antitumor activity induced by Toxoplasma, mice were sacrificed weekly and blood, muscle, lung, and spleen tissues were collected. In addition, we checked the antiangiogenic activity and cytotoxic T-lymphocyte responses one week after infection with T. gondii. Untreated age- and sex-matched control mice were treated with equivalent doses of normal saline.
In vivo evaluation of antitumor activity
According to the experimental protocol, mice were injected with LLC cells, T. gondii parasites or Quil-A. LLC cells were implanted at a density of 1×105 viable cells into the femoral muscle and Quil-A (20 µg per mouse) was injected intraperitoneally twice weekly for three consecutive weeks. Mice were orally infected with five cysts of the T. gondii Me49 strain. The extent of tumor growth was measured weekly using sterile metric calipers. Tumor volume was calculated using the following formula (13): tumor volume (µL)=tumor width (mm)2×tumor length (mm)×0.5.
Histopathological analysis
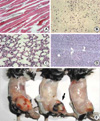
The lung and muscle samples from each mouse were removed immediately after anesthesia and placed in 10% buffered neutral formaldehyde (Polyscience Inc., Warrington, PA, U.S.A.). Paraffin-embedded tissues were cut and stained with hematoxylin and eosin (H-E), and the cancer cells were examined using a bright-field microscope.
Enzyme-linked immunosorbent assay (ELISA)
Serum samples were obtained from each mouse, and the IgG subclasses were quantified. TLA was prepared according to the protocol outlined by Lee et al. (15). Each well of a 96-well plate was coated with TLA (10 µg/mL) and incubated overnight at 4℃. After blocking, serum samples were diluted 1:100 in 0.1% bovine serum albumin/phosphate-buffered saline (BSA/PBS) that contained 0.05% Tween-20, and 100 µL of sample was added to each well. The plates were incubated for 2 hr, and HRP-conjugated goat anti-mouse IgG1 or IgG2a (Southern Biotechnology Associates Inc., Birmingham, AL, U.S.A.) was added. Following the final washing step, freshly prepared O-phenylenediamine dihydrochloride was added, and the reaction was stopped by the addition of 4 N H2SO4. The optical density was read at 492 nm using an automated ELISA reader (Tecan A-5082, Salzburg, Austria).
Splenocyte preparation and flow cytometry
Following removal, the spleens were homogenized and erythrocytes were lysed in Tris-NH4Cl (pH 7.2). Splenocyte extracts that contained 1×106 cells were incubated with 50 µL of fluorescein isothiocyanate (FITC)-conjugated anti-mouse CD4+ and CD8+ monoclonal antibodies or an isotype-specific control (1:100 dilution in 0.1% BSA/PBS; PharMingen, San Diego, CA, U.S.A.) for 1 hr at 4℃. The cells were washed three times in 0.1% BSA/PBS with centrifugation, and then fixed with 1% paraformaldehyde and analyzed by flow cytometry (FACScan; Becton Dickinson, Franklin Lakes, NJ, U.S.A.). All data were analyzed using the Cell-Quest program (Becton Dickinson).
RT-PCR assessment of cytokines
Splenic cytokine mRNA expression was assessed according to the methods of Lee et al. (15). Each sample was processed for total mRNA isolation using the RNAgent kit (Promega, Madison, WI, U.S.A.). The cDNA was prepared using a starting mixture that contained 5 µg of total RNA, 8 µL of 5×RT buffer, 4 µL of dNTPs, 25 µM oligo (dT), and 1 µL of avian myeloblastosis virus (AMV) reverse transcriptase. PCR was performed using 2-13 µL of the cDNA reaction mixture that contained 10×polymerase buffer, 250 µM dNTPs, 0.4 µM of the 3'-primer and 5'-primer, and 2.5 U Taq polymerase. Each PCR cycle consisted of 94℃ for 1 min, 57℃ for 1 min, and 72℃ for 1 min. The cytokine primer sequences were as follows: for hypoxanthine phosporibosyltransferase (HPRT), 3'-GAGGGTAGGCTGGCCTATGGCT-5' and 5'-TTGGATACAGGCCAGACTTTGTTG-3'; for interferon gamma (IFN-γ), 3'-CTCATGGAATGCATCCTTTTTCG-5' and 5'-ACGCTACACAC-TGCATCTTGG-3'; and for interleukin-10 (IL-10), 3'-TGTCTAGGTCCTGGAGTCCAGCAGACTCAA-5' and 5'-CCAGTTTTACCTGGTAGAAGTGATG-3'. After 35 PCR cycles, the products were separated by electrophoresis on 2% agarose gels. Quantification of mRNA was performed with an imaging densitometer (Bio-Rad, Hercules, CA, U.S.A.).
Cytotoxic T lymphocyte assay
One week after T. gondii (or cancer cell) injection, splenic lymphocytes were isolated from each group and cultured in RPMI 1640 for three days. The splenocytes were then co-cultured with irradiated LLC cells at various effector to target (E:T) ratios for seven days. One µCi of 51Cr (NEN Research Products, Boston, MA, U.S.A.) was added to each well and the plate was allowed to incubate for 3 hr. The plate was then washed several times with RPMI 1640 until the culture supernatant contained <500 counts per minute (CPM) of 51Cr. The plates were incubated for an additional 4 hr at 37℃, followed by removal of 100 µL of each sample, and the CPM were determined by a gamma counter (Beckman, Fullerton, CA, U.S.A.). The percent lysis was calculated as follows: percent lysis=(mean CPM of test sample-mean CPM of spontaneous release)/(mean CPM of maximal release-mean CPM of spontaneous release)×100.
Matrigel angiogenesis assay
Matrigel angiogenesis assay was performed as described by Passaniti et al. (16). One week after T. gondii (or cancer cell) injection, the C57BL/6 mice were injected subcutaneously near the abdominal mid-line using a 25G needle with 0.5 mL Matrigel (BD Bioscience, Bedford, MA, U.S.A.) that contained 300 ng of basic fibroblast growth factor (bFGF; Sigma Chemical Co., St. Louis, MO, U.S.A.) and 32 U/mL heparin (Sigma). The injected Matrigel rapidly formed a single, solid gel and the mice were sacrificed six days later. The underlying skin of the mouse was removed, and the gels were recovered for hemoglobin (Hb) measurement using the Drabkin reagent kit (Sigma). Hemoglobin concentrations were calculated from a known Hb control, which was run in parallel.
Statistical analysis
All results are expressed as the mean±standard error (SE) for each group. Kaplan-Meier and Log-Rank tests were performed to analyze the differences in survival rates. To evaluate differences in tumor sizes, antibody titers, phenotype profiles, cytokine mRNA levels, Hb concentrations, and cytotoxicity, Kruskal-Wallis tests were used to check for statistical differences among the groups. Differences between groups were considered significant at p<0.05.
RESULTS
Increased survival rates of cancer-bearing mice by T. gondii infection
Mice were treated with T. gondii parasites, LLC cells, or Quil-A according to our protocol, and survival rates were evaluated over eight weeks. As shown in Fig. 1, the TG-, QA-, and TG/QA-injected mice survived the entire experimental period. However, the LLC- and LLC/QA-injected mice began dying 28 days after cancer cell injection, and all of these mice died after 39 days. The survival rate of the TG/LLC-injected mice was 20%, and the TG/LLC/QA-injected mice had a 30% survival rate at 8 weeks after cancer cell injection, which were significantly higher than the rates for the LLC- and LLC/QA-injected mice (p<0.05).
T. gondii infection inhibits the tumor growth in mice
Mice infected with T. gondii developed brain cysts within two weeks of infection (data not shown). Cancer cells were observed in the muscle tissues within one week in the LLC- and LLC/QA-injected groups (Fig. 2A, B). However, cancer cells were found in the TG/LLC- and TG/LLC/QA-injected mice by two weeks. In addition, cancer cells were found in the lungs three weeks after LLC cell injection in the LLC- and LLC/QA-injected groups (Fig. 2C, D), whereas cancer they were found within five weeks in TG/LLC- and TG/LLC/QA-injected mice. The skin overlying the tumor began to ulcerate three to four weeks after cancer cell injection in the LLC-injected mice (Fig. 2E), whereas ulceration was observed within five weeks in the TG/LLC- and TG/LLC/QA-injected mice.
Tumor masses were palpable by two to three weeks after cancer cell injection in the cancer-bearing LLC-, LLC/QA-, TG/LLC-, and TG/LLC/QA-injected mice. In the LLC- and LLC/QA-injected mice, tumor masses increased abruptly in size at two to four weeks after cancer cell injection, and the tumor volumes at four weeks were 8,760±465 and 6,740±360 µL, respectively (Fig. 3). The tumor volumes were significantly decreased in the TG/LLC- and TG/LLC/QA-injected mice (2,155±520 µL and 1,980±342 µL, respectively) at four weeks as compared with the LLC- and LLC/QA-injected mice (p<0.05).
IgG2a titers were increased in T. gondii-infected cancer-bearing mice
Serum samples were assayed for IgG subclasses against TLA by ELISA. The specific IgG1 and IgG2a titers of the LLC-, QA-, and LLC/QA-injected mice were similar to those of control mice. However, the specific IgG1 titers of the TG- and TG/QA-injected mice were significantly increased after one week of T. gondii infection (Fig. 4A). In addition, the IgG1 titers of the TG/LLC- and TG/LLC/QA-injected mice were markedly depressed following cancer cell injection as compared with the TG- and TG/QA-injected mice (p<0.05).
The IgG2a titers of the TG- and TG/QA-injected mice were significantly increased after T. gondii infection as compared to the control mice. In the TG/LLC- and TG/LLC/QA-injected mice, the IgG2a titers following cancer cell injection were significantly depressed until four weeks compared with those of the TG- and TG/QA-injected mice, although the IgG2a titers increased abruptly thereafter (Fig. 4B).
CD4+ and CD8+ T-cells are decreased in cancer-bearing mice
To evaluate changes in T-cell subtypes following either T. gondii infection or cancer cell injection, we analyzed the phenotypic profiles of murine splenocytes. The percentages of CD4+ T-cells in the TG-injected mice were temporarily decreased at one to two weeks post-infection compared to those of the control mice, and these levels returned to baseline shortly thereafter (Table 1). The percentages of CD4+ T-cells in the LLC-, LLC/QA-, TG/LLC-, and TG/LLC/QA-injected mice were significantly decreased following cancer cell injection and remained markedly depressed after three weeks (p<0.05).
As shown in Table 2, the percentages of CD8+ T-cells in the LLC-, LLC/QA-, TG/LLC-, and TG/LLC/QA-injected mice were significantly decreased three weeks after cancer cell injection. The LLC- and LLC/QA-injected mice maintained these low values, whereas the percentage of CD8+ T-cells in the TG/LLC- and TG/LLC/QA-injected mice recovered slightly at five weeks after T. gondii infection.
IFN-γ mRNA expression was significantly increased in T. gondii-infected cancer-bearing mice
As shown in Fig. 5A, the levels of IFN-γ mRNA expression in the LLC- and LLC/QA-injected mice were increased two- to three-fold at three weeks after cancer cell injection as compared to the control (p<0.05). However, the levels of IFN-γ mRNA expression in the TG-, TG/QA-, and TG/LLC/QA-injected mice were increased immediately after T. gondii infection, peaked two to three weeks post-infection (14-19-fold increase), and slowly decreased thereafter. However, IFN-γ mRNA expression in the TG/LLC-injected mice was significantly lower than that in the TG-, TG/QA-, and TG/LLC/QA-injected mice, even though IFN-γ mRNA expression in the TG/LLC-injected mice was significantly increased by one week post-infection.
As compared to the control mice, the levels of IL-10 mRNA expression in the LLC- and LLC/QA-injected mice increased significantly, by 1.3-2.3-fold, three weeks after cancer cell injection. The levels of IL-10 mRNA expression in the TG-, TG/QA-, and TG/LLC-injected mice increased immediately following T. gondii infection, peaking at three weeks post-infection (5.5-7.5-fold increase). However, IL-10 mRNA expression in the TG/LLC/QA-injected mice, which was significantly lower than in the TG-, TG/QA-, and TG/LLC-injected mice from 3 weeks postinfection, peaked at two weeks and subsequently decreased.
T. gondii infection induced tumor cytolysis by splenocytes
An in vitro CTL assay was performed to determine whether the splenic T-cells of Toxoplasma-infected cancer-bearing mice increased cytotoxic activity against LLC cells. As shown in Fig. 6, the LLC-, QA-, and LLC/QA-injected mice did not show CTL responses. However, the splenocyte CTL activities of the TG- and TG/QA-injected mice increased as the E:T ratio increased. The percentages of specific lysis seen in the TG/LLC- and TG/LLC/QA-injected mice were considerably higher than those in the LLC- and LLC/QA-injected mice. However, the CTL activities of the TG/LLC- and TG/LLC/QA-injected mice were significantly lower than those of the TG- and TG/QA-injected mice (p<0.05).
T. gondii infection inhibits tumor angiogenesis in vivo
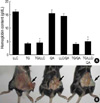
The Matrigel plugs that formed following subcutaneous implantation of Matrigel alone produced little or no local reaction, nor did they produce an angiogenic response. However, the plugs that formed following the implantation of Matrigel that was supplemented with bFGF and heparin caused an angiogenic reaction (Fig. 7). We measured the Hb concentrations inside the Matrigel plugs in order to quantify the extent of angiogenesis. The Hb concentrations in the LLC-, QA-, and LLC/QA-injected mice ranged from 14.6±2.0 to 16.2±1.6 g/dL, whereas those in the TG- and TG/QA-injected mice ranged from 4.1±0.7 to 4.3±0.9 g/dL. The Hb concentrations in the TG/LLC- and TG/LLC/QA-injected mice were significantly lower than those in the LLC- or LLC/QA-injected mice (p<0.05), but were not significantly different from those in the TG- or TG/QA-injected mice.
DISCUSSION
Tumors frequently interfere with the development and function of immune responses (3-5). The ability of various infections to suppress neoplastic growth has been well documented. In addition, T. gondii is a nonspecific immune stimulant used for cancer immunotherapy (7-11). In the present study, we examined the mechanisms of the antitumor effects of T. gondii infection on lung cancer in vivo. Toxoplasma infection of LLC-bearing mice significantly increased the survival rates, serum IgG2a titers, IFN-γ mRNA expression, CD8+ T-cell percentages, CTL responses, and also inhibited angiogenesis. These values were similar or further improved by the addition of an adjuvant, Quil-A. These results indicate that T. gondii activates Th1 immune responses and inhibits angiogenesis in LLC-bearing mice, leading to the induction of antitumor and antimetastatic activities.
Cancer immunotherapy attempts to harness the exquisite power and specificity of the immune system for the treatment of malignancy. Although cancer cells are less immunogenic than pathogens, the immune system is clearly capable of recognizing and eliminating tumor cells. Lung cancer immunotherapy may represent one new approach that has low toxicity and high specificity, but implementation has been a challenge due to poor antigenic characterization and the ability of lung cancers to escape immune responses (3-5, 17). Several different immunotherapeutic treatment strategies and mouse models have been developed. In this study, we utilized LLC cells, which remain highly tumorigenic in C57BL/6 mice and produce primary tumors and lung metastases histologically indistinguishable from the original tumor line (18). In the LLC- and LLC/QA-injected mice, tumor masses were initially palpable in the muscle tissues located at the site of cancer cell injection from two weeks after cancer cell injection, and cancer cells were found in the lungs three weeks after LLC cell injection. Thus, it was confirmed that the LLC cell line is useful for studying the mechanisms of cancer chemotherapeutic agents. After T. gondii infection, tumor volumes were significantly decreased in the TG/LLC- and TG/LLC/QA-injected mice as compared with the LLC- and LLC/QA-injected mice. However, the mechanisms of antitumor activity induced by T. gondii infection are not easily introduced in LLC, so we suggested the mechanisms of antitumor effects in this study.
Infection with T. gondii results in a strong cell-mediated immune response, especially the T-cell-mediated response (19). In the present study, the percentages of CD8+ and CD4+ T-cells were significantly decreased in cancer-bearing LLC- and LLC/QA-injected mice. Following infection with T. gondii, the percentages of CD8+ T-cells in TG/LLC- or TG/LLC/QA-injected mice were increased from 5 weeks post-infection, but those of CD4+ T-cells were not. Collectively, these data indicate that T-cell-mediated immune responses were severely suppressed in Lewis lung carcinoma, and T. gondii alone or in combination with Quil-A may provide an appropriate epitope for MHC-1 presentation (17, 19), and therefore, the percentages of CD8+ T-cells were recovered. This phenomenon is confirmed by the results of a cytotoxic T lymphocyte assay. Namely, Toxoplasma-infected TG/LLC- and TG/LLC/QA-injected mice demonstrated higher CTL responses of CD8+ T-cells than the cancer-bearing LLC- or LLC/QA-injected mice. Previous studies have also demonstrated that strong antitumor responses are induced by either abundant infiltration of both CD4+ and CD8+ T-cells and tumor-specific CTLs in LLC-bearing mice after treatment with IL-2 and IL-12 (17) or increases in the numbers of apoptotic tumor cells, CD8+ T-cells, and natural killer (NK) cells that invade the tumors after injection with ergosterol (14).
Cytokines serve as critical regulators of cell-mediated immunity by both enhancing and limiting immune responses (20). The CD4+ T-cells are divided into two classes on the basis of the range of cytokines produced. Th1 cells produce IL-2 and IFN-γ, and control the production of IgG2a, whereas Th2 cells produce IL-4, IL-5, and IL-10, and control the production of IgG1 and IgE. Because of these interleukin secretion patterns, Th1 cells promote cell-mediated immunity. In contrast, Th2 cells induce B-cell differentiation and inhibit cell-mediated immunity and antitumor responses. In the present study, we found that IFN-γ mRNA expression in TG/LLC-injected mice was significantly higher than that in LLC-injected mice and significantly lower than that in TG-injected mice. These results indicate that Th1 immune responses were depressed in Lewis lung carcinoma, and show that the production of cytokines such as IFN-γ, TNF-α, IL-12, and IL-2 can arrest proliferation of malignant cells and prevent the angiogenesis necessary for tumor growth (3, 5, 17, 21, 22). Similar results were reported in melanoma, liver cancer, and thyroid cancer (22-24). And these phenomena were also confirmed in the titrations of the IgG subclasses. The IgG2a titer, which is the primary indicator of a Th1 immune response (25), was significantly increased in the TG/LLC- and TG/LLC/QA-injected mice four weeks after infection, while the IgG1 titer, an indicator of the Th2 immune response, remained unchanged.
The development of a vascular supply is essential for the growth, maturation, and maintenance of normal tissues. It is also required for wound healing and rapid growth of solid tumors, and is involved in various other pathologic conditions. Tumor angiogenesis is regulated by the production of angiogenic stimulators, including members of the fibroblast growth factor and the vascular endothelial growth factor families (13, 14, 26, 27). In the present study, a combination of bFGF and heparin recruited vessels from the surrounding tissues into the Matrigel. The Hb concentrations in T. gondii-infected TG-, TG/QA-, TG/LLC-, and TG/LLC/QA-injected mice were significantly decreased by three- to four-fold as compared to uninfected QA-, LLC-, and LLC/QA-injected mice. In a similar study, Hunter et al. (28) showed that T. gondii infection is accompanied by strong systemic suppression of angiogenesis. This suppression results in severe hypoxia and avascular necrosis, which are incompatible with progressive neoplastic growth. Beside this, heyneanol A obtained from an Oriental medicinal herb, sodium pyroglutamate isolated from Agaricus blazei, and glycosaminoglycan isolated from the African giant snail were also reported as potent antitumor and anti-angiogenic substances in LLC-bearing mice (13, 14, 26, 27).
The antitumor activity of cancer immunotherapy has been explored using advanced molecular and immunological methods. Host immunity is likely related to various factors, including the route of challenge, mouse strain, antigen preparation, and adjuvant (29). Our present results suggest that Toxoplasma infection in LLC-bearing mice induces antitumor activity against lung cancer through the induction of a Th1-type immune response and antiangiogenic activity. These effects were similar or further increased by the modulation of Quil-A. Quil-A, a saponin derived from the bark of Quillaia saponaria Molina, induces antigen presentation, cytokine production, and T-lymphocyte cytotoxicity (29, 30). However, the molecular mechanisms underlying Quil-A activity have not yet been completely described. Infection with T. gondii leads to the rapid induction of a strong cell-mediated immune response in the experimental murine host. These facts suggest the possibility that new immunostimulating molecules could be found in Toxoplasma organisms.







 XML Download
XML Download