

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Ceramides are the main lipids in the stratum corneum, and their depletion is thought to be one of the etiological factors for barrier disruption in various levels of skin conditions (1-4). Marked depletion of ceramides in the stratum corneum has been reported especially in patients with psoriasis and atopic dermatitis.
Ceramides are generated by serine palmitoyltransferase (SPT), the enzyme for de novo synthesis or by the action of sphingomyelinase. As lipid second messengers, they mediate antiproliferative and apoptotic effect via activation of several signal transduction molecules such as protein kinase C-alpha (PKC-α) and c-jun N-terminal kinase (JNK) (5-7). In our previous study, the decreased levels of ceramide in psoriatic skin lesions were shown to be responsible for epidermal hyperplasa via downregulation of proapoptotic signal cascade such as PKC-α and JNK. In contrast, diacyglycerol (DAG) levels were elevated and free fatty acid levels were not changed (8, 9). However, in the past study, the ceramide level and protein expression of ceramide-metabolizing enzymes in psoriatic skin lesion was not elucidated. In the present study measured the expression of SPT, key enzyme for de novo ceramide synthesis, and ceramidase, the degradative enzyme of ceramide, in both psoriatic epidermis and non-lesional epidermis.
MATERIALS AND METHODS
Patients and skin biopsies
Twenty four Korean patients with psoriasis (9 women, 15 men) ranging in age from 9 to 37 yr gave informed consent and took part in this study. All the subjects had psoriasis vulgaris as identified through clinical and histological assessment and had not been treated either systemically or topically for at least 1 month before punch biopsies were done (Table 1). Using a 4-mm punch, biopsies were taken from lesional and non-lesional skin on the lower extremities, back, or arms. The epidermis was separated as described previously (10). Specifically, the epidermis was separated from whole-skin biopsies by overnight incubation at 4℃ in a 1:1 (v/v) mixture of dispase solution (Roche Molecular Biochemicals, Manheim, Germany) and Hank's balanced salt solution (HBSS; Gibco BRL, Life Technologies, Rockville, MD, U.S.A.).
Assessing the clinical severity of psoriasis
The clinical severity was assessed using the psoriasis area and severity index (PASI) score, which is calculated as follows: PASI=0.1 (Eh+Ih+Dh)Ah+0.2 (Eu+Iu+Du)Au+0.3 (Et+It+Dt)At+0.4 (El+Il+Dl)Al, where E=erythema, I=infiltration, D=desquamation, A=area, h=head, u=upper extremities, t=trunk, and l=lower extremities. A numerical value is given to the extent of the lesions in each area: 1=<10%, 2=10-30%, 3=30-50%, 4=50-70%, 5=70-90%, and 6=90-100%. E, I, and D were scored on a five-point scale (0=no symptoms, 1=slight, 2=moderate, 3=marked, and 4=very marked) to obtain a final PASI score between 0 and 72. The PASI scores of the patients who took part in this study ranged between 2.0 and 27.4. The patients with PASI scores <30 were enrolled in this study in order to determine whether alterations in the levels of SPT and ceramidase are closely correlated to the clinical severity in psoriatic patients (Table 1). All patients with PASI scores >30 were excluded in this study because their skin lesions were shown to be transformed into severe variants of psoriasis, such as the erythrodermic type.
Western blot analysis
Every frozen skin sample was added to 400 µL of Folch solution (CHCl3: MeOH 2:1, v/v mixture) and was homogenized using a polytron homogenizer, and 200 µL of 0.1 M KCl was added. The mixture was centrifuged at 3,000 rpm for 5 min each. The upper phase containing the extracted proteins was separated. This process was repeated one more time. The protein concentration was determined by BCA protein assay (Pierce, Rockford, U.S.A.) according to the manufacturer's protocol. Fifty micrograms of protein was fractionated on 10% Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis (SDS-PAGE) and transferred to a nitrocellulose membrane by electrophoresis. The membranes were blocked with 5% non-fat dry milk for 1 hr at room temperature and incubated with prepared human SPT, ceramidase, and tubulin antibody (Labfrontier, Seoul, Korea) at a 1:250 dilution with Tris-buffered saline containing 0.05% Tween-20 (TSB-T) for 1 hr. After washing with TBS-T for 1 hr, the membranes were reacted with horseradish peroxidase-conjugated anti-rabbit IgG antibody (Amersham Biosciences, Buckinghamshire, U.K.) diluted 1:250 with TBS-T for 1 hr at room temperature. After washing the membranes with TBS-T for 1 hr, the protein level was detected using the ECL chemiluminescence kit (Amersham Bioscience, Buckinghamshire, U.K.).
Statistical analysis
The statistical significance of the difference in the levels of SPT/tubulin and ceramidase/tubulin between the lesional and the non-lesional samples of psoriasis patients was assessed using a paired t-test. The correlations between the levels of SPT and the PASI scores were determined using the Spearman correction test. p values less than 0.05 were considered statistically significant.
RESULTS
Protein expression of Serine palmitoyltransferase and ceramidase in psoriatic epidermis and non-lesional epidermis
Western blot analysis of SPT and ceramidase in the lesional and non-lesional epidermis of psoriasis patients is shown in Fig. 1, 2. When the level of SPT protein expression was quantified by densitometry and was expressed as a ratio of SPT to tubulin (Fig. 1, 2), the levels of SPT expression in the lesional epidermis was significantly lower (range, 1.26-5.59) than those in the non-lesional epidermis ( range, 4.63-9.97) (Fig. 1). In contrast, levels of ceramidase expression in the lesional epidermis ranged from 0.025 to 1.070, and those in the non-lesional epidermis ranged from 0.023 to 1.556, which indicates that there was no significant difference in the level of ceramidase of lesional and non-lesional epidermis of psoriasis (Fig. 2). After normalization with tubulin, the mean level of SPT/tubuin in the lesional epidermis was significantly decreased compared to the non-lesional epidermis as shown in Fig. 3A (p<0.05). However, there was no significant difference in the mean ratio of ceramidase to tubulin between lesional psoriatic skin and non-lesional skin (Fig. 3B).
Correlation of SPT, ceramidase, and PASI score
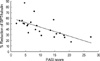
The protein expression of SPT and ceramidase of non-lesion and lesion, and the PASI score in psoriatic patients are shown in Table 1. Although the absolute level of SPT varied from patient to patient, SPT expression of lesional epidermis was significantly less (p<0.05) than that of non-lesional epidermis, as with the percent reduction ranging from 17.5 to 87.6. When the PASI score was applied, there was a significant negative correlation between percent reduction of SPT expression in the lesional epidermis and the PASI score (r=-0.76, p=0.01) as shown in Fig. 4. However, there was no significant correlation between percent reduction of ceramidase expression in the lesional epidermis and the PASI score.
DISCUSSION
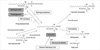
The epidermis contains a large amount of sphingolipids, which are concentrated in upper, differentiated cell layer, where they are thought to participate in the maintenance of the cutaneous permeability barrier (11). Among the epidermal sphingolipids, ceramide is the main lipid in the stratum corneum and its depletion is thought to be one of the etiological factors for barrier disruption in skin conditions (1-4). Moreover, as a signal transduction molecule, ceramide is involved in the cell proliferation and apoptosis signaling pathway via activation of several signal transduction molecules, and its depletion is related with various skin disease such as atopic dermatitis or psoriasis (5-8). Synthesis of ceramide is regulated by several enzymes such as sphingomyelinase, β-glucocerebrosidase, and ceramidase (12-15). Also a de novo pathway mediated by SPT functions as a key pathway of ceramide systhesis (16) (Fig. 5).
SPT is a key enzyme in sphingolipid biosynthesisis. It is the first committed and rate-limiting step in sphigolipid synthesis (17, 18). One of the substrates for SPT is various acyl-CoAs. In the mammalian cells, palmitoyl CoA is one of the most abundant acly-CoA types and it is the predominant acyl-CoA substrate of SPT in vivo (19-21). However, SPT strictly utilizes L-serine as its amino acid substate (22). In human epidermis, SPT catalyzes the palmitoyl CoA and L-serine to synthesize ceramide (17).
Ceramidase hydrolyzes ceramide into shpingophosphorylcholine and free fatty acid. It is demonstrated that marked elevated activity of this enzyme is responsible for low levels of cermaide in stratum corneum of atopic dermatitis patients (23).
In our previous study, decreased ceramide in psoriatic skin lesions is shown to be responsible for epidermal hyperplasia via downregulation of proapoptotic signal cascade such as PKC-α and JNK. In contrast, DAG levels were elevated and free fatty acid levels were not changed (8, 9). However, in the past study, we could not elucidate what makes ceramide decreased in psoriatic skin lesion.
In this study, we measured the expression of SPT and ceramidase in both psoriatic epidermis and non-lesional epidermis. SPT is one of the enzymes, which is related to the synthesis of ceramide, and ceramidase is a catalytic enzyme, which is related to the decrease of the levels of ceramide. The results of this experiment showed that levels of SPT were significantly decreased in the psoriatic skin. However, the levels of ceramidase showed no significant difference between psoriatic epidermis and non-lesional epidermis. We also evaluated the relationship between the PASI score and decreased levels of SPT to determine the role of decreased SPT on the severity of psoriasis. The correlation between the percentage reduction in the ratio of SPT/tubulin in the lesional epidermis and the PASI scores showed a significantly negative correlation. Considering above results, decreased levels of ceramide in psoriatic skin, as shown in our previous study, are responsible for decreased levels of SPT in psoriatic skin lesion, and their levels of reduction are highly related to the clinical severity of psoriasis. However, according to this experiment, ceramidase does not seem to have a relation with psoriatic epidermal changes, and this result is different from that of the previous study on ceramidase activity in atopic dermatitis (23).
In conclusion, we demonstrated that the decreased levels of ceramide in the psoriatic epidermis is related to its de novo synthetic pathway, which is associated with levels of SPT, and the levels of SPT are closely related to the clinical severity of psoriasis. Considering that psoriasis is one of the inflammatory skin diseases, several cytokines or growth factors secreted by keratinocytes, Langerhans cells, or T-cells in the psoriatic skin lesion seem to affect the expression of serine palmitoyltransferase, and this postulation need further studies.


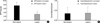

 XML Download
XML Download